

Characterization of germination receptors of Bacillus cereus ATCC 14579
- PMID: 16391023
- PMCID: PMC1352193
- DOI: 10.1128/AEM.72.1.44-53.2006
Characterization of germination receptors of Bacillus cereus ATCC 14579
Abstract
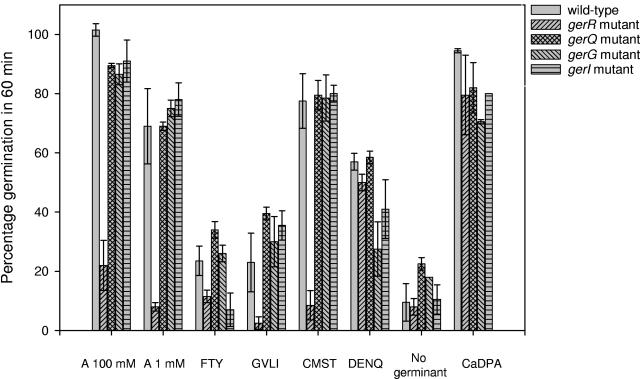
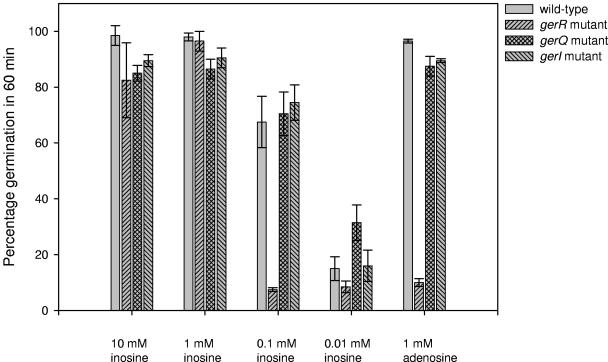
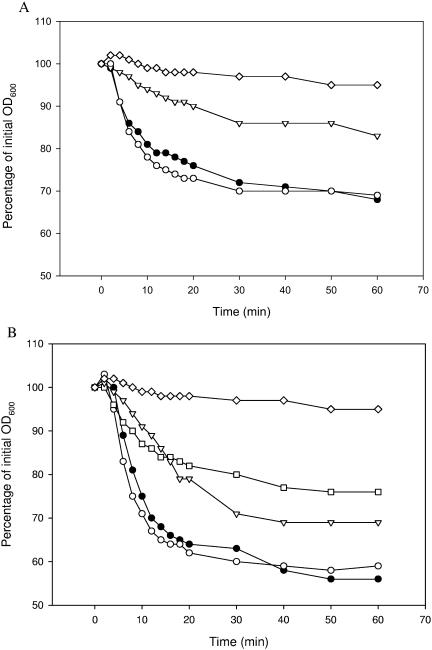
Specific amino acids, purine ribonucleosides, or a combination of the two is required for efficient germination of endospores of Bacillus cereus ATCC 14579. A survey including 20 different amino acids showed that l-alanine, l-cysteine, l-threonine, and l-glutamine are capable of initiating the germination of endospores of B. cereus ATCC 14579. In addition, the purine ribonucleosides inosine and adenosine can trigger germination of the spores. Advanced annotation of the B. cereus ATCC 14579 genome revealed the presence of seven putative germination (ger) operons, termed gerG, gerI, gerK, gerL, gerQ, gerR, and gerS. To determine the role of the encoded putative receptors in nutrient-induced germination, disruption mutants were constructed by the insertion of pMUTIN4 into each of the seven operons. Four of the seven mutants were affected in the germination response to amino acids or purine ribonucleosides, whereas no phenotype could be attributed to the mutants with disrupted gerK, gerL, and gerS loci. The strain with a disrupted gerR operon was severely hampered in the ability to germinate: germination occurred in response to l-glutamine but not in the presence of any of the other amino acids tested. The gerG mutant showed significantly reduced l-glutamine-induced germination, which points to a role of this receptor in the l-glutamine germination signaling pathway. gerR, gerI, and gerQ mutants showed reduced germination rates in the presence of inosine, suggesting a role for these operons in ribonucleoside signaling. Efficient germination by the combination of l-glutamine and inosine was shown to involve the gerG and gerI operons, since the germination of mutants lacking either one of these receptors was significantly reduced. Germination triggered by the combination of l-phenylalanine and inosine was lost in the gerI mutant, indicating that both molecules are effective at the GerI receptor.
Figures

Similar articles
-
gerR, a novel ger operon involved in L-alanine- and inosine-initiated germination of Bacillus cereus ATCC 14579.Appl Environ Microbiol. 2005 Feb;71(2):774-81. doi: 10.1128/AEM.71.2.774-781.2005. Appl Environ Microbiol. 2005. PMID: 15691930 Free PMC article.
-
Germination of Bacillus cereus spores in response to L-alanine and to inosine: the roles of gerL and gerQ operons.Microbiology (Reading). 2002 Jul;148(Pt 7):2089-2095. doi: 10.1099/00221287-148-7-2089. Microbiology (Reading). 2002. PMID: 12101297
-
Germinant receptor diversity and germination responses of four strains of the Bacillus cereus group.Int J Food Microbiol. 2010 Apr 30;139(1-2):108-15. doi: 10.1016/j.ijfoodmicro.2010.01.028. Epub 2010 Jan 25. Int J Food Microbiol. 2010. PMID: 20153067
-
The Ger receptor family from sporulating bacteria.Curr Issues Mol Biol. 2010;12(3):147-58. Curr Issues Mol Biol. 2010. PMID: 20472940 Free PMC article. Review.
-
Symposium on immunosuppressive drugs. Comparative actions of some purine analogues on growing Bacillus cereus.Fed Proc. 1967 May-Jun;26(3):905-6. Fed Proc. 1967. PMID: 4960737 Review. No abstract available.
Cited by
-
The Cooperative and Interdependent Roles of GerA, GerK, and Ynd in Germination of Bacillus licheniformis Spores.Appl Environ Microbiol. 2016 Jun 30;82(14):4279-4287. doi: 10.1128/AEM.00594-16. Print 2016 Jul 15. Appl Environ Microbiol. 2016. PMID: 27208128 Free PMC article.
-
Dissecting interactions between nucleosides and germination receptors in Bacillus cereus 569 spores.Microbiology (Reading). 2010 Apr;156(Pt 4):1244-1255. doi: 10.1099/mic.0.030270-0. Epub 2009 Dec 24. Microbiology (Reading). 2010. PMID: 20035009 Free PMC article.
-
Influence of Sporulation Temperature on Germination and Growth of B. weihenstephanensis Strains in Specific Nutrients and in an Extended Shelf-Life Refrigerated Matrix Under Commercial Pasteurization and Storage Conditions.Foods. 2024 Oct 28;13(21):3434. doi: 10.3390/foods13213434. Foods. 2024. PMID: 39517218 Free PMC article.
-
Kinetic evidence for the presence of putative germination receptors in Clostridium difficile spores.J Bacteriol. 2010 Aug;192(16):4215-22. doi: 10.1128/JB.00488-10. Epub 2010 Jun 18. J Bacteriol. 2010. PMID: 20562307 Free PMC article.
-
Investigating the functional hierarchy of Bacillus megaterium PV361 spore germinant receptors.J Bacteriol. 2013 Jul;195(13):3045-53. doi: 10.1128/JB.00325-13. Epub 2013 Apr 26. J Bacteriol. 2013. PMID: 23625848 Free PMC article.
References
-
- Barlass, P. J., C. W. Houston, M. O. Clements, and A. Moir. 2002. Germination of Bacillus cereus spores in response to l-alanine and to inosine: the roles of gerL and gerQ operons. Microbiology 148:2089-2095. - PubMed
-
- Corfe, B. M., R. L. Sammons, D. A. Smith, and C. Mauel. 1994. The gerB region of the Bacillus subtilis 168 chromosome encodes a homologue of the gerA spore germination operon. Microbiology 140:471-478. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
