

Protease inhibitors fail to prevent pore formation by the activated Bacillus thuringiensis toxin Cry1Aa in insect brush border membrane vesicles
- PMID: 16391085
- PMCID: PMC1352293
- DOI: 10.1128/AEM.72.1.506-515.2006
Protease inhibitors fail to prevent pore formation by the activated Bacillus thuringiensis toxin Cry1Aa in insect brush border membrane vesicles
Abstract
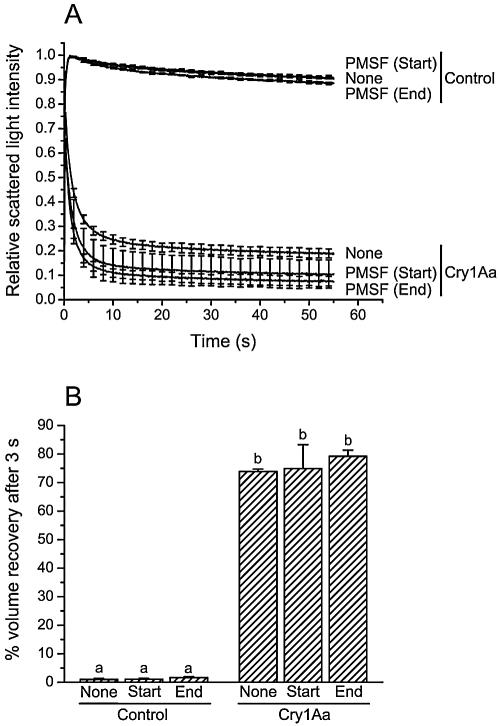
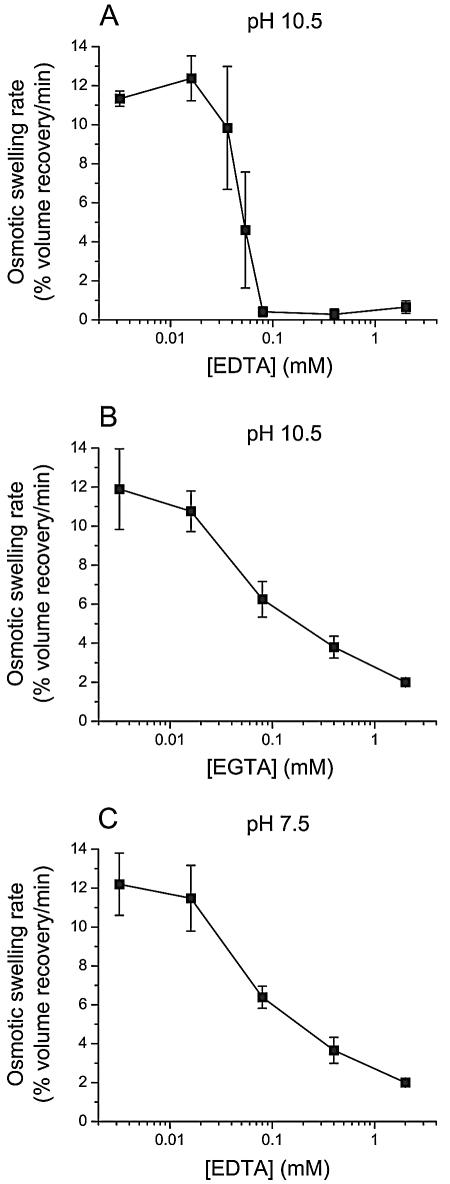
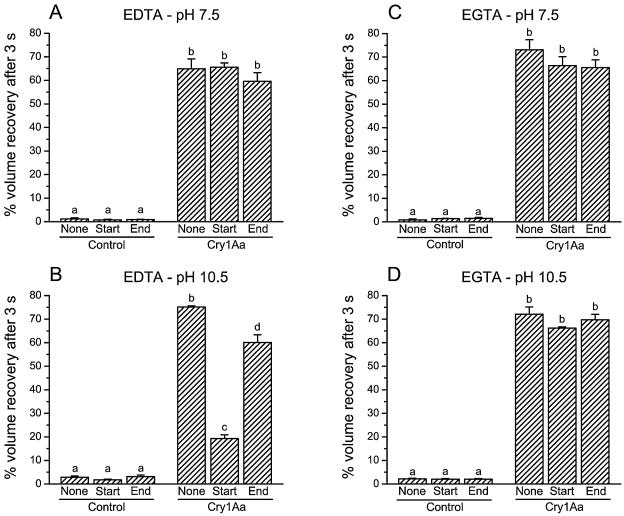
To investigate whether membrane proteases are involved in the activity of Bacillus thuringiensis insecticidal toxins, the rate of pore formation by trypsin-activated Cry1Aa was monitored in the presence of a variety of protease inhibitors with Manduca sexta midgut brush border membrane vesicles and by a light-scattering assay. Most of the inhibitors tested had no effect on the pore-forming ability of the toxin. However, phenylmethylsulfonyl fluoride, a serine protease inhibitor, promoted pore formation, although this stimulation only occurred at higher inhibitor concentrations than those commonly used to inhibit proteases. Among the metalloprotease inhibitors, o-phenanthroline had no significant effect; EDTA and EGTA reduced the rate of pore formation at pH 10.5, but only EDTA was inhibitory at pH 7.5. Neither chelator affected the properties of the pores already formed after incubation of the vesicles with the toxin. Taken together, these results indicate that, once activated, Cry1Aa is completely functional and does not require further proteolysis. The effect of EDTA and EGTA is probably better explained by their ability to chelate divalent cations that could be necessary for the stability of the toxin's receptors or involved elsewhere in the mechanism of pore formation.
Figures
Similar articles
-
Mutations in domain I interhelical loops affect the rate of pore formation by the Bacillus thuringiensis Cry1Aa toxin in insect midgut brush border membrane vesicles.Appl Environ Microbiol. 2009 Jun;75(12):3842-50. doi: 10.1128/AEM.02924-08. Epub 2009 Apr 17. Appl Environ Microbiol. 2009. PMID: 19376918 Free PMC article.
-
Helix 4 mutants of the Bacillus thuringiensis insecticidal toxin Cry1Aa display altered pore-forming abilities.Appl Environ Microbiol. 2004 Oct;70(10):6123-30. doi: 10.1128/AEM.70.10.6123-6130.2004. Appl Environ Microbiol. 2004. PMID: 15466558 Free PMC article.
-
Role of helix 3 in pore formation by the Bacillus thuringiensis insecticidal toxin Cry1Aa.Biochemistry. 2002 May 14;41(19):6178-84. doi: 10.1021/bi011572e. Biochemistry. 2002. PMID: 11994014
-
N-terminal activation is an essential early step in the mechanism of action of the Bacillus thuringiensis Cry1Ac insecticidal toxin.J Biol Chem. 2002 Jul 5;277(27):23985-7. doi: 10.1074/jbc.C200263200. Epub 2002 May 17. J Biol Chem. 2002. PMID: 12019259
-
Insect midgut structures and molecules as targets of plant-derived protease inhibitors and lectins.Pest Manag Sci. 2019 May;75(5):1212-1222. doi: 10.1002/ps.5233. Epub 2018 Nov 16. Pest Manag Sci. 2019. PMID: 30306668 Review.
Cited by
-
Bt toxin modification for enhanced efficacy.Toxins (Basel). 2014 Oct 22;6(10):3005-27. doi: 10.3390/toxins6103005. Toxins (Basel). 2014. PMID: 25340556 Free PMC article. Review.
-
Characterization of vegetative insecticidal protein vip genes of Bacillus thuringiensis from Sichuan Basin in China.Curr Microbiol. 2011 Mar;62(3):752-7. doi: 10.1007/s00284-010-9782-3. Epub 2010 Oct 21. Curr Microbiol. 2011. PMID: 20963416
-
Candidalysin Is the Hemolytic Factor of Candida albicans.Toxins (Basel). 2022 Dec 15;14(12):874. doi: 10.3390/toxins14120874. Toxins (Basel). 2022. PMID: 36548771 Free PMC article.
-
Fate of transgenic DNA from orally administered Bt MON810 maize and effects on immune response and growth in pigs.PLoS One. 2011;6(11):e27177. doi: 10.1371/journal.pone.0027177. Epub 2011 Nov 23. PLoS One. 2011. PMID: 22132091 Free PMC article.
-
Genetic transformation and pyramiding of aprotinin-expressing sugarcane with cry1Ab for shoot borer (Chilo infuscatellus) resistance.Plant Cell Rep. 2010 Apr;29(4):383-95. doi: 10.1007/s00299-010-0829-5. Epub 2010 Feb 24. Plant Cell Rep. 2010. PMID: 20179936
References
-
- Angsuthanasombat, C., N. Crickmore, and D. J. Ellar. 1993. Effects on toxicity of eliminating a cleavage site in a predicted interhelical loop in Bacillus thuringiensis CryIVB δ-endotoxin. FEMS Microbiol. Lett. 111:255-262. - PubMed
-
- Bah, A., K. van Frankenhuyzen, R. Brousseau, and L. Masson. 2004. The Bacillus thuringiensis Cry1Aa toxin: effects of trypsin and chymotrypsin site mutations on toxicity and stability. J. Invertebr. Pathol. 85:120-127. - PubMed
-
- Barrett, A. J. 1994. Classification of peptidases. Methods Enzymol. 244:1-15. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
