

The Thermotoga maritima phenotype is impacted by syntrophic interaction with Methanococcus jannaschii in hyperthermophilic coculture
- PMID: 16391122
- PMCID: PMC1352257
- DOI: 10.1128/AEM.72.1.811-818.2006
The Thermotoga maritima phenotype is impacted by syntrophic interaction with Methanococcus jannaschii in hyperthermophilic coculture
Abstract
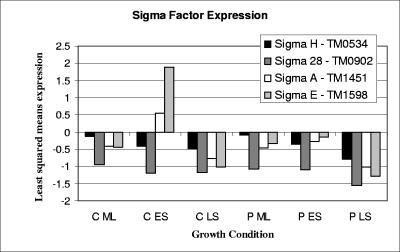
Significant growth phase-dependent differences were noted in the transcriptome of the hyperthermophilic bacterium Thermotoga maritima when it was cocultured with the hyperthermophilic archaeon Methanococcus jannaschii. For the mid-log-to-early-stationary-phase transition of a T. maritima monoculture, 24 genes (1.3% of the genome) were differentially expressed twofold or more. In contrast, methanogenic coculture gave rise to 292 genes differentially expressed in T. maritima at this level (15.5% of the genome) for the same growth phase transition. Interspecies H2 transfer resulted in three- to fivefold-higher T. maritima cell densities than in the monoculture, with concomitant formation of exopolysaccharide (EPS)-based cell aggregates. Differential expression of specific sigma factors and genes related to the ppGpp-dependent stringent response suggests involvement in the transition into stationary phase and aggregate formation. Cell aggregation was growth phase dependent, such that it was most prominent during mid-log phase and decayed as cells entered stationary phase. The reduction in cell aggregation was coincidental with down-regulation of genes encoding EPS-forming glycosyltranferases and up-regulation of genes encoding beta-specific glycosyl hydrolases; the latter were presumably involved in hydrolysis of beta-linked EPS to release cells from aggregates. Detachment of aggregates may facilitate colonization of new locations in natural environments where T. maritima coexists with other organisms. Taken together, these results demonstrate that syntrophic interactions can impact the transcriptome of heterotrophs in methanogenic coculture, and this factor should be considered in examining the microbial ecology in anaerobic environments.
Figures

References
-
- Adams, M. W. W. 1990. The metabolism of hydrogen by extremely thermophilic, sulfur-dependent archaebacteria. FEMS Microbiol. Rev. 75:219-238.
-
- Allison, D. G., B. Ruiz, C. SanJose, A. Jaspe, and P. Gilbert. 1998. Extracellular products as mediators of the formation and detachment of Pseudomonas fluorescens biofilms. FEMS Microbiol. Lett. 167:179-184. - PubMed
-
- Bibel, M., C. Brettl, U. Gosslar, G. Kriegshauser, and W. Liebl. 1998. Isolation and analysis of genes for amylolytic enzymes of the hyperthermophilic bacterium Thermotoga maritima. FEMS Microbiol. Lett. 158:9-15. - PubMed
-
- Bonch-Osmolovskaya, E. A., and K. O. Stetter. 1991. Interspecies hydrogen transfer in cocultures of thermophilic Archaea. Syst. Appl. Microbiol. 14:205-208.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
