

Olfactory nerve-evoked, metabotropic glutamate receptor-mediated synaptic responses in rat olfactory bulb mitral cells
- PMID: 16394070
- PMCID: PMC2366052
- DOI: 10.1152/jn.01150.2005
Olfactory nerve-evoked, metabotropic glutamate receptor-mediated synaptic responses in rat olfactory bulb mitral cells
Abstract
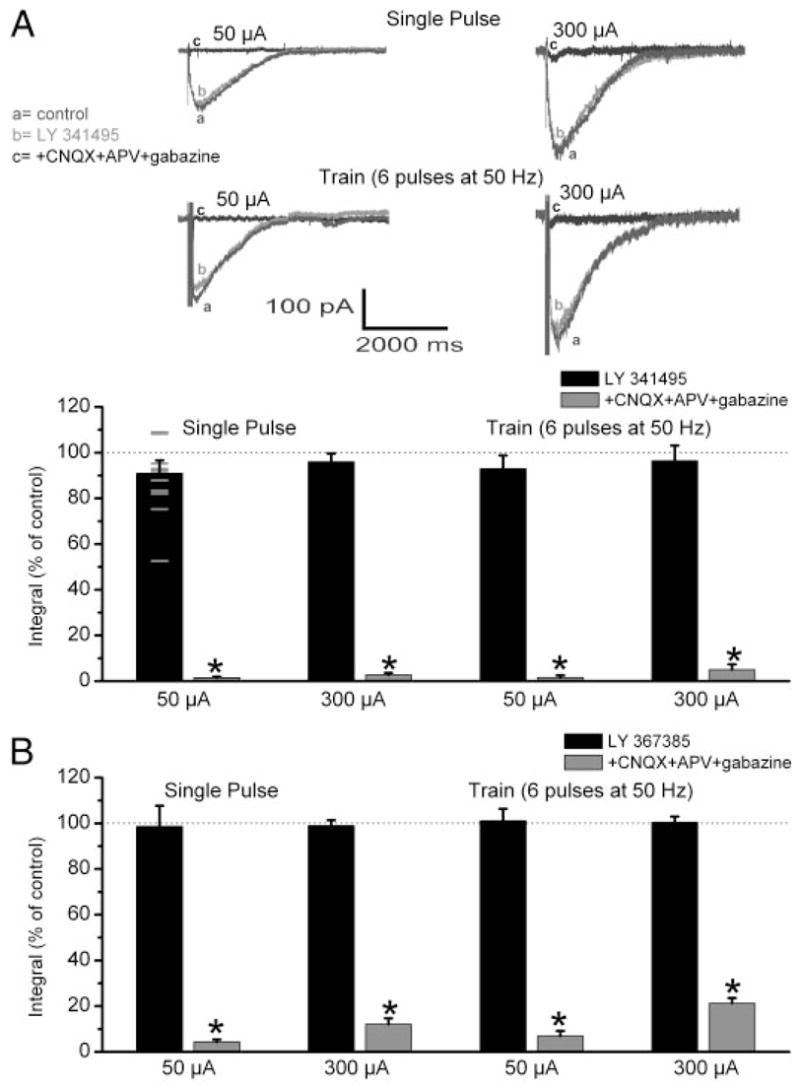
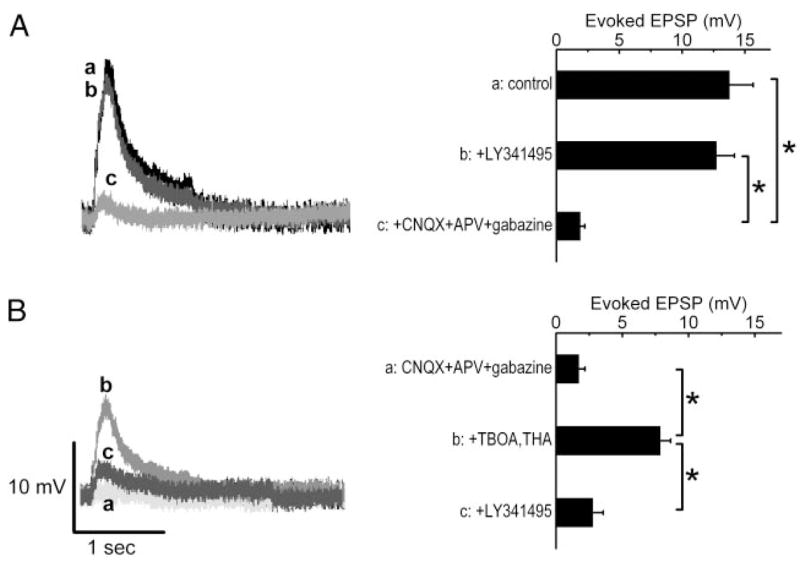
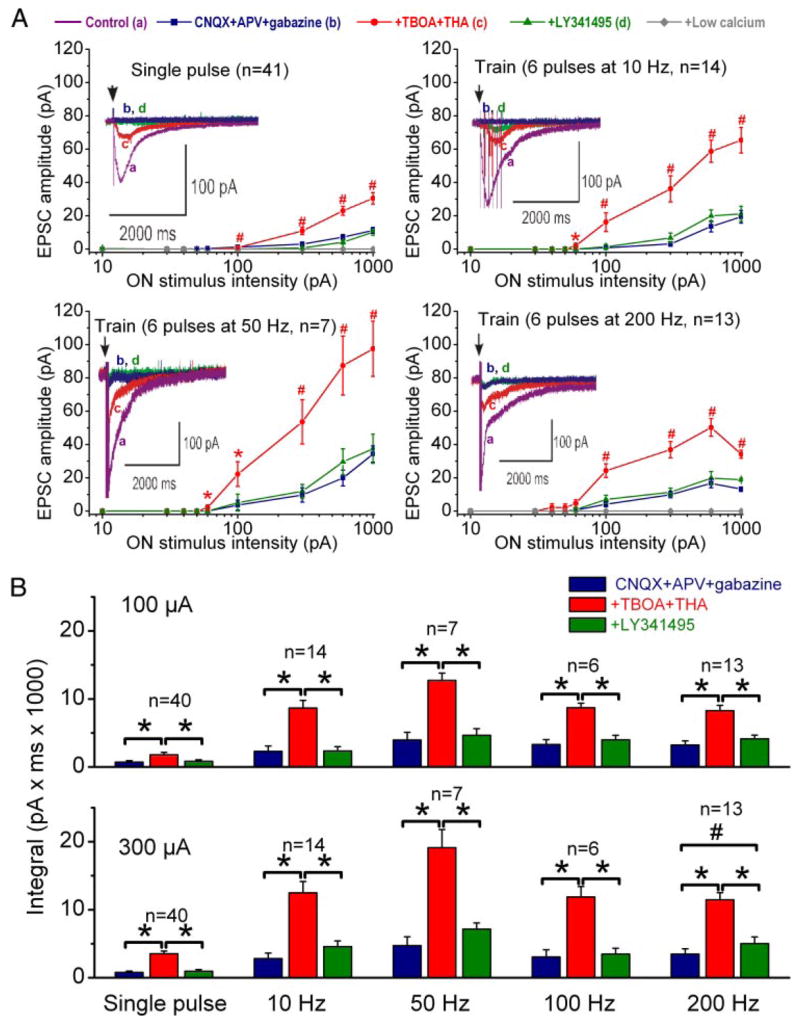
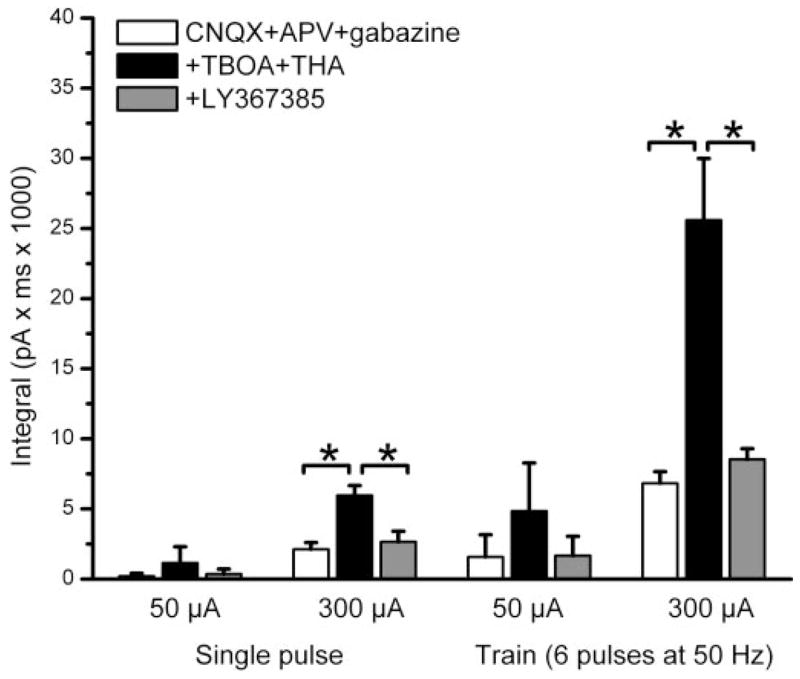
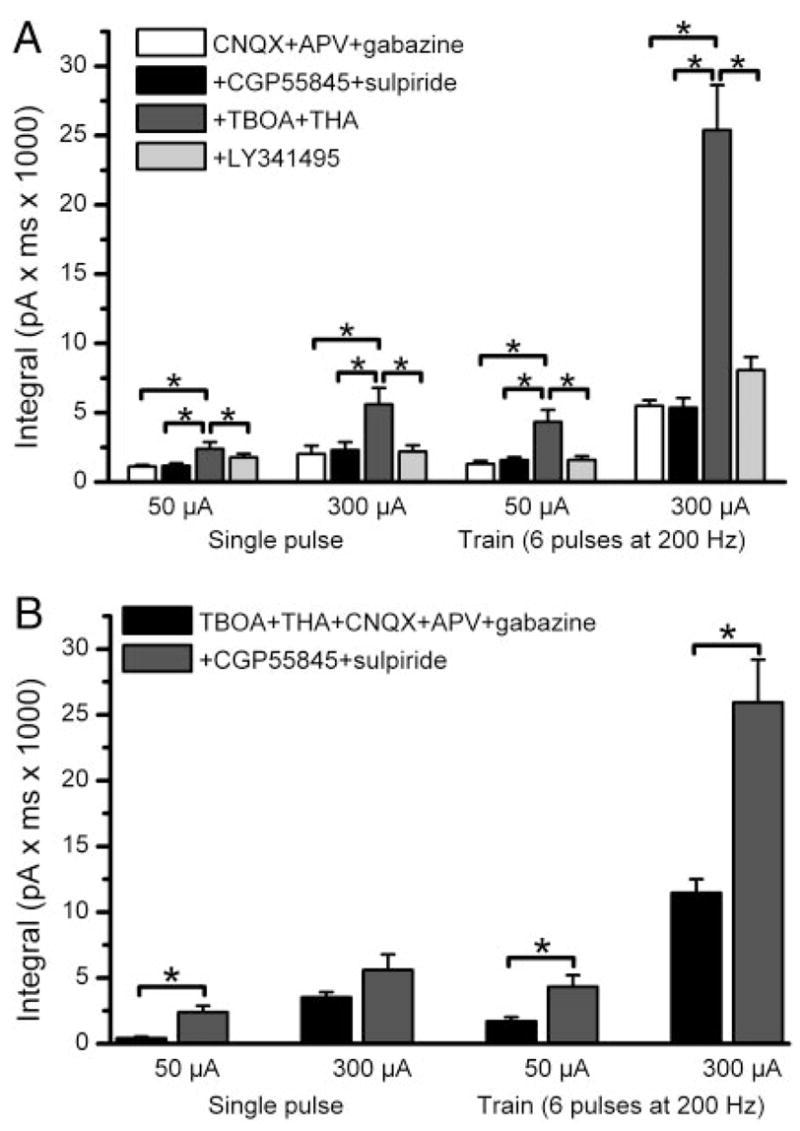
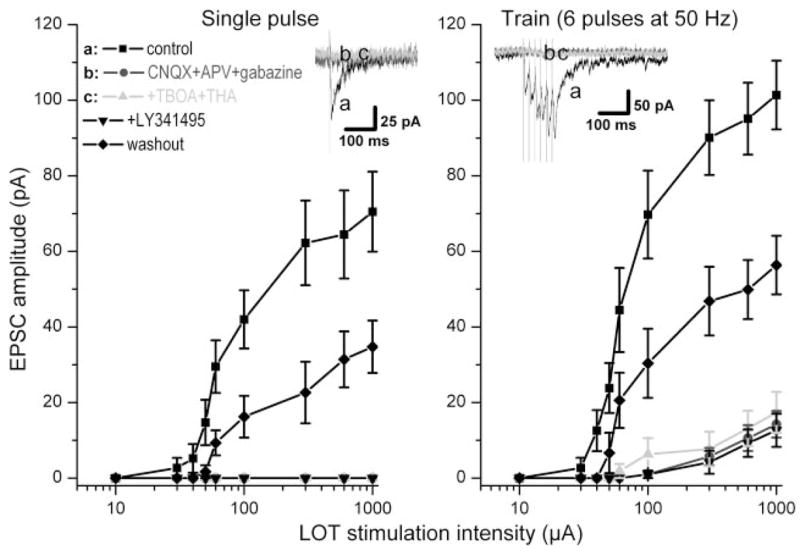
The group I metabotropic glutamate receptor (mGluR) subtype, mGluR1, is highly expressed on the apical dendrites of olfactory bulb mitral cells and thus may be activated by glutamate released from olfactory nerve (ON) terminals. Previous studies have shown that mGluR1 agonists directly excite mitral cells. In the present study, we investigated the involvement of mGluR1 in ON-evoked responses in mitral cells in rat olfactory bulb slices using patch-clamp electrophysiology. In voltage-clamp recordings, the average EPSC evoked by single ON shocks or brief trains of ON stimulation (six pulses at 50 Hz) in normal physiological conditions were not significantly affected by the nonselective mGluR antagonist LY341495 (50-100 microM) or the mGluR1-specific antagonist LY367385 (100 microM); ON-evoked responses were attenuated, however, in a subset (36%) of cells. In the presence of blockers of ionotropic glutamate and GABA receptors, application of the glutamate uptake inhibitors THA (300 microM) and TBOA (100 microM) revealed large-amplitude, long-duration responses to ON stimulation, whereas responses elicited by antidromic activation of mitral/tufted cells were unaffected. Magnitudes of the ON-evoked responses elicited in the presence of THA-TBOA were dependent on stimulation intensity and frequency, and were maximal during high-frequency (50-Hz) bursts of ON spikes, which occur during odor stimulation. ON-evoked responses elicited in the presence of THA-TBOA were significantly reduced or completely blocked by LY341495 or LY367385 (100 microM). These results demonstrate that glutamate transporters tightly regulate access of synaptically evoked glutamate from ON terminals to postsynaptic mGluR1s on mitral cell apical dendrites. Taken together with other findings, the present results suggest that mGluR1s may not play a major role in phasic responses to ON input, but instead may play an important role in shaping slow oscillatory activity in mitral cells and/or activity-dependent regulation of plasticity at ON-mitral cell synapses.
Figures
Similar articles
-
Tonic and synaptically evoked presynaptic inhibition of sensory input to the rat olfactory bulb via GABA(B) heteroreceptors.J Neurophysiol. 2000 Sep;84(3):1194-203. doi: 10.1152/jn.2000.84.3.1194. J Neurophysiol. 2000. PMID: 10979995
-
Regulation of main olfactory bulb mitral cell excitability by metabotropic glutamate receptor mGluR1.J Neurophysiol. 2004 Nov;92(5):3085-96. doi: 10.1152/jn.00349.2004. Epub 2004 Jun 22. J Neurophysiol. 2004. PMID: 15212418
-
Metabotropic glutamate receptors in the main olfactory bulb drive granule cell-mediated inhibition.J Neurophysiol. 2007 Jan;97(1):858-70. doi: 10.1152/jn.00884.2006. Epub 2006 Nov 8. J Neurophysiol. 2007. PMID: 17093122 Free PMC article.
-
Dopamine D2 receptor-mediated presynaptic inhibition of olfactory nerve terminals.J Neurophysiol. 2001 Dec;86(6):2986-97. doi: 10.1152/jn.2001.86.6.2986. J Neurophysiol. 2001. PMID: 11731555
-
Olfactory nerve stimulation-evoked mGluR1 slow potentials, oscillations, and calcium signaling in mouse olfactory bulb mitral cells.J Neurophysiol. 2006 May;95(5):3097-104. doi: 10.1152/jn.00001.2006. Epub 2006 Feb 8. J Neurophysiol. 2006. PMID: 16467433
Cited by
-
Group I mGluR activation enhances Ca(2+)-dependent nonselective cation currents and rhythmic bursting in main olfactory bulb external tufted cells.J Neurosci. 2009 Sep 23;29(38):11943-53. doi: 10.1523/JNEUROSCI.0206-09.2009. J Neurosci. 2009. PMID: 19776280 Free PMC article.
-
Dissecting the signaling mechanisms underlying recognition and preference of food odors.J Neurosci. 2014 Jul 9;34(28):9389-403. doi: 10.1523/JNEUROSCI.0012-14.2014. J Neurosci. 2014. PMID: 25009271 Free PMC article.
-
Insulin-like growth factor 1 regulates excitatory synaptic transmission in pyramidal neurons from adult prefrontal cortex.Neuropharmacology. 2022 Oct 1;217:109204. doi: 10.1016/j.neuropharm.2022.109204. Epub 2022 Aug 2. Neuropharmacology. 2022. PMID: 35931212 Free PMC article.
-
Ultrastructural Contributions to Extrasynaptic Glutamatergic Signaling in Olfactory Bulb Glomeruli.J Comp Neurol. 2025 Mar;533(3):e70034. doi: 10.1002/cne.70034. J Comp Neurol. 2025. PMID: 40090882
-
Cellular and Synaptic Mechanisms That Differentiate Mitral Cells and Superficial Tufted Cells Into Parallel Output Channels in the Olfactory Bulb.Front Cell Neurosci. 2020 Dec 22;14:614377. doi: 10.3389/fncel.2020.614377. eCollection 2020. Front Cell Neurosci. 2020. PMID: 33414707 Free PMC article.
References
-
- Aroniadou-Anderjaska V, Ennis M, Shipley MT. Current-source density analysis in the rat olfactory bulb: laminar distribution of kainate/AMPA and NMDA receptor-mediated currents. J Neurophysiol. 1999a;81:15–28. - PubMed
-
- Aroniadou-Anderjaska V, Ennis M, Shipley MT. Dendrodendritic recurrent excitation in mitral cells of the rat olfactory bulb. J Neurophysiol. 1999b;82:489–494. - PubMed
-
- Aroniadou-Anderjaska V, Zhou FM, Priest CA, Ennis M, Shipley MT. Tonic and synaptically evoked presynaptic inhibition of sensory input to the rat olfactory bulb via GABA(B) heteroreceptors. J Neurophysiol. 2000;84:1194–1203. - PubMed
-
- Aroniadou-Anderjaska VA, Ennis M, Shipley MT. Glomerular synaptic responses to olfactory nerve input in rat olfactory bulb slices. Neuroscience. 1997;79:425–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous