

DnaA couples DNA replication and the expression of two cell cycle master regulators
- PMID: 16395331
- PMCID: PMC1383511
- DOI: 10.1038/sj.emboj.7600927
DnaA couples DNA replication and the expression of two cell cycle master regulators
Abstract
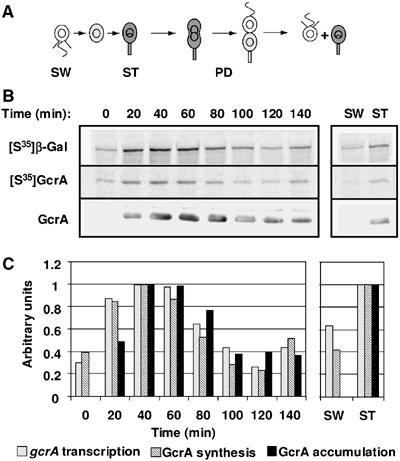
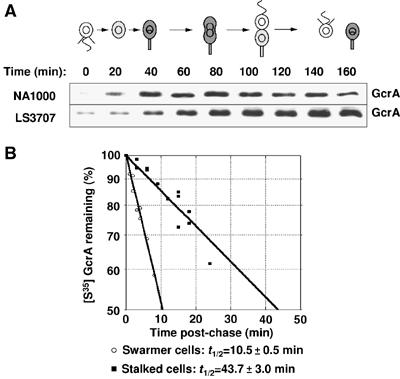
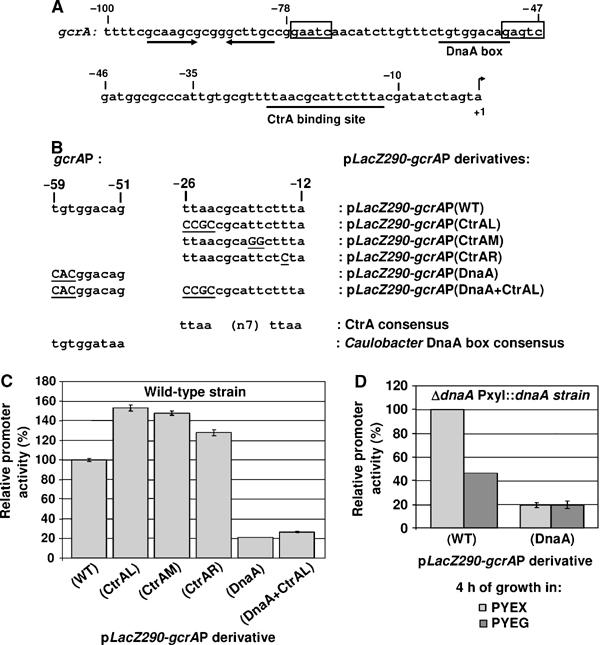
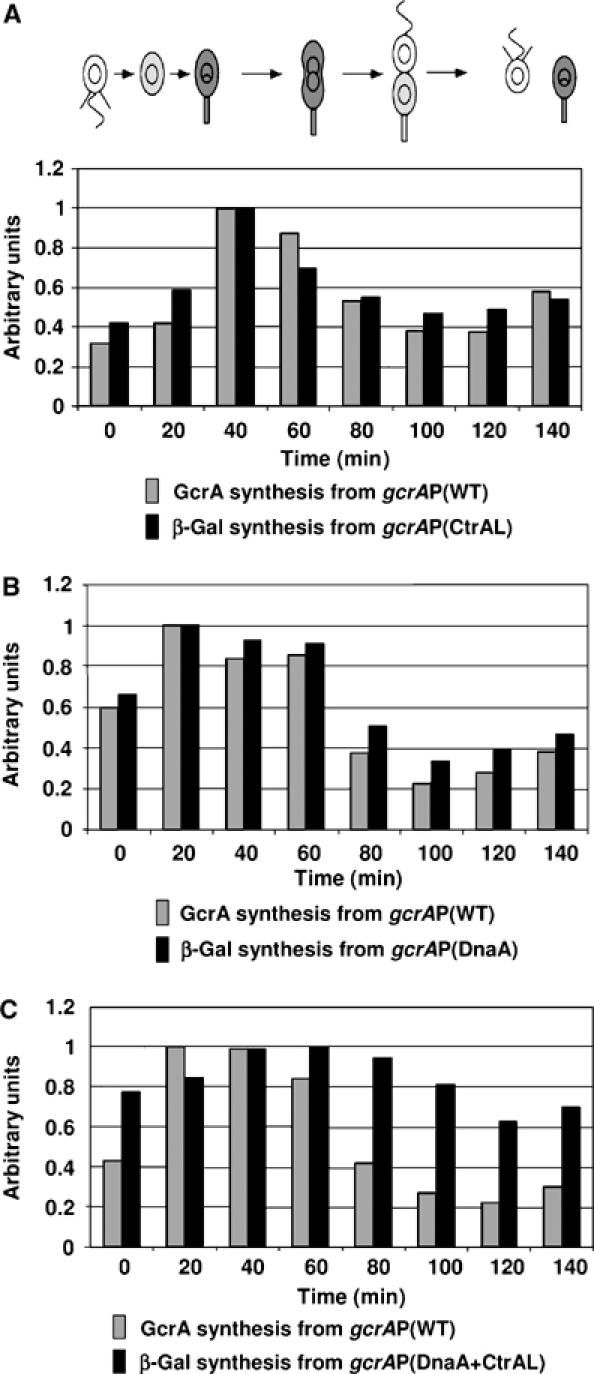
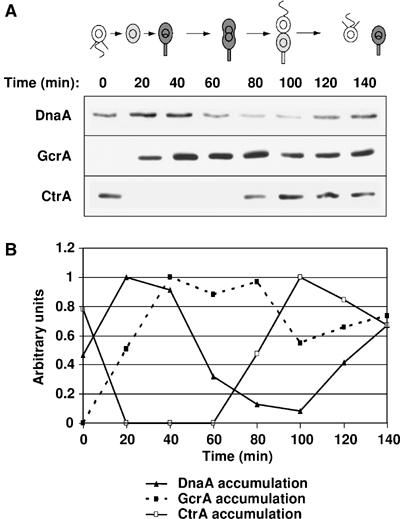
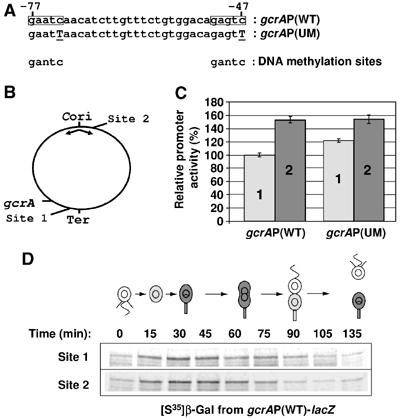
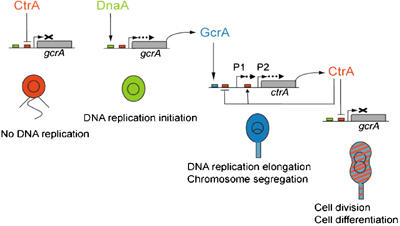
Cell cycle progression in Caulobacter is driven by the master transcriptional regulators CtrA and GcrA. The cellular levels of CtrA and GcrA are temporally and spatially out-of-phase during the cell cycle, with CtrA repressing gcrA transcription and GcrA activating ctrA transcription. Here, we show that DnaA, a protein required for the initiation of DNA replication, also functions as a transcriptional activator of gcrA, which in turn activates multiple genes, notably those involved in chromosome replication and segregation. The cellular concentration of DnaA is cell cycle-controlled, peaking at the time of replication initiation and gcrA induction. Regulated proteolysis of GcrA contributes to the cell cycle variations in GcrA abundance. We propose that DnaA couples DNA replication initiation with the expression of the two oscillating regulators GcrA and CtrA and that the DnaA/GcrA/CtrA regulatory cascade drives the forward progression of the Caulobacter cell cycle.
Figures
References
-
- Bramhill D, Kornberg A (1988) Duplex opening by DnaA protein at novel sequences in initiation of replication at the origin of the E. coli chromosome. Cell 52: 743–755 - PubMed
-
- Domian IJ, Quon KC, Shapiro L (1997) Cell type-specific phosphorylation and proteolysis of a transcriptional regulator controls the G1-to-S transition in a bacterial cell cycle. Cell 90: 415–424 - PubMed
-
- Fuller RS, Funnell BE, Kornberg A (1984) The DnaA protein complex with the E. coli chromosomal replication origin (oriC) and other DNA sites. Cell 38: 889–900 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
