

lessen encodes a zebrafish trap100 required for enteric nervous system development
- PMID: 16396911
- PMCID: PMC2651469
- DOI: 10.1242/dev.02215
lessen encodes a zebrafish trap100 required for enteric nervous system development
Abstract
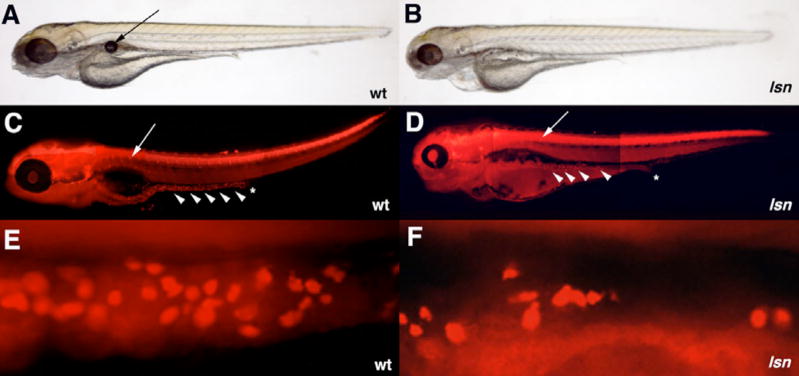
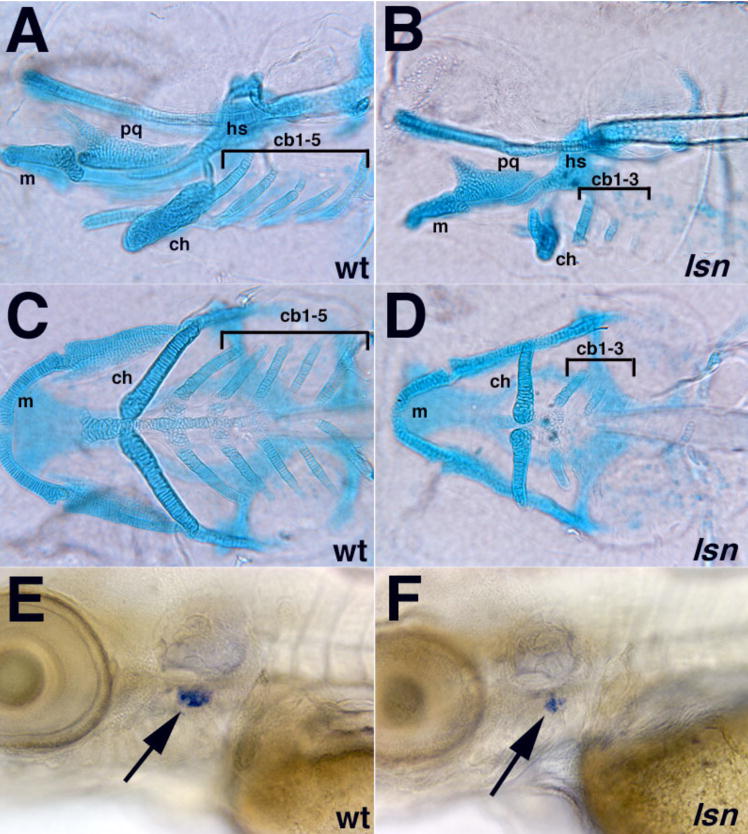
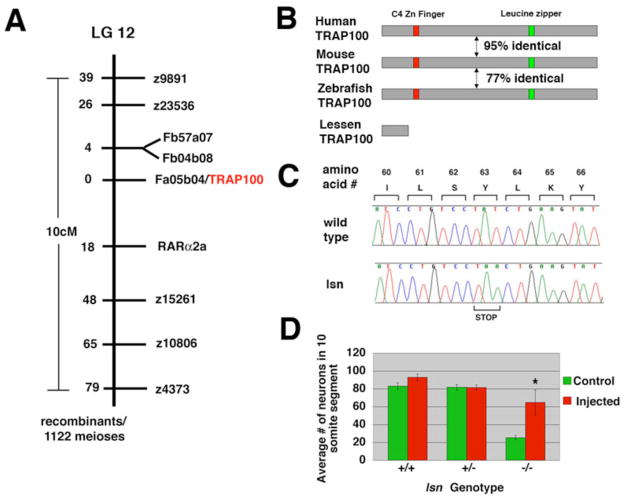
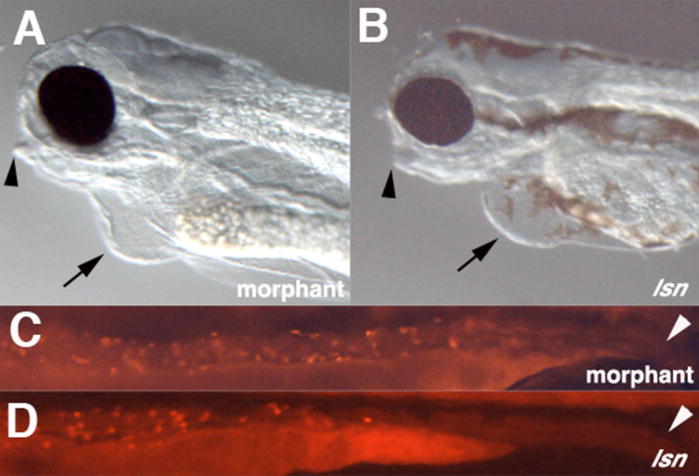
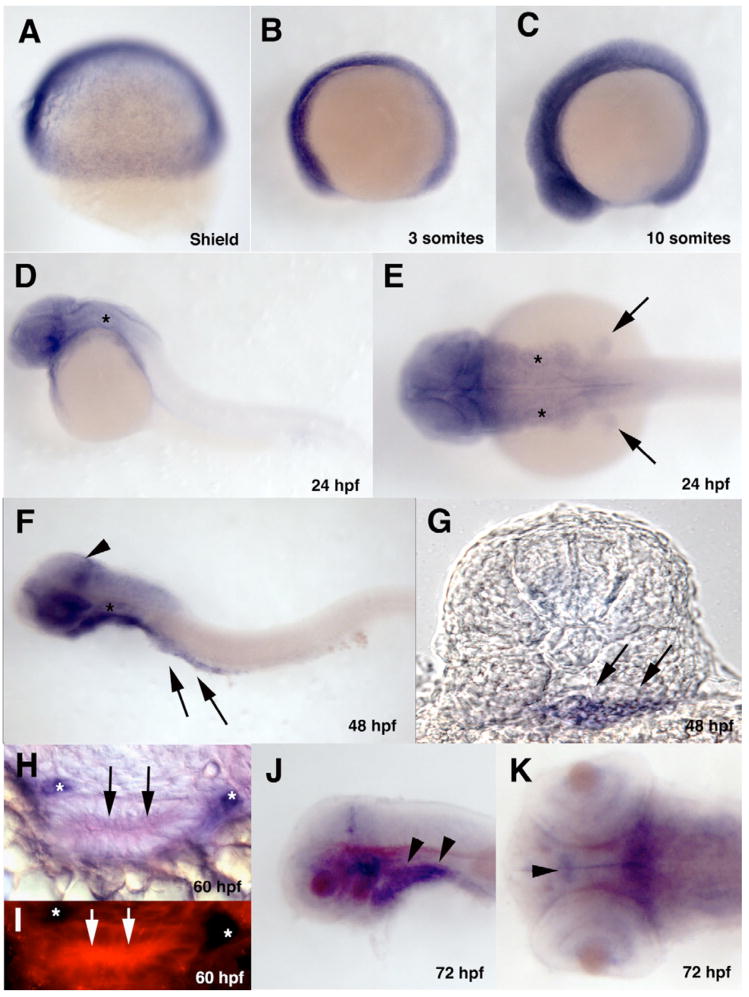
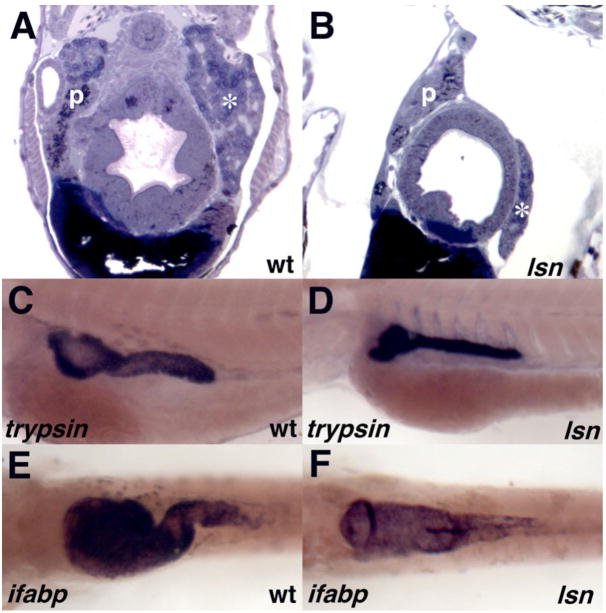
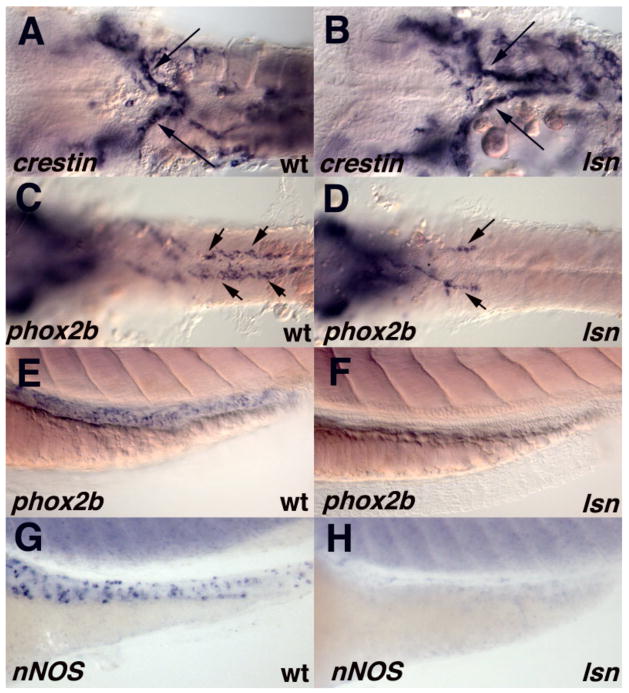
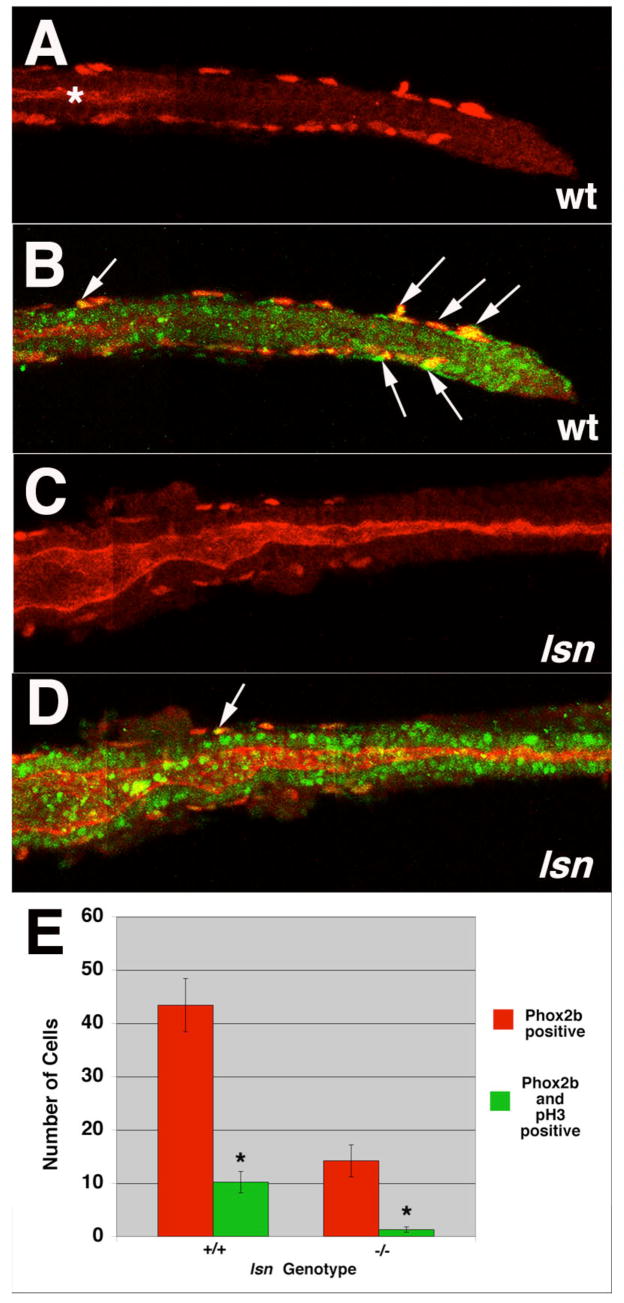
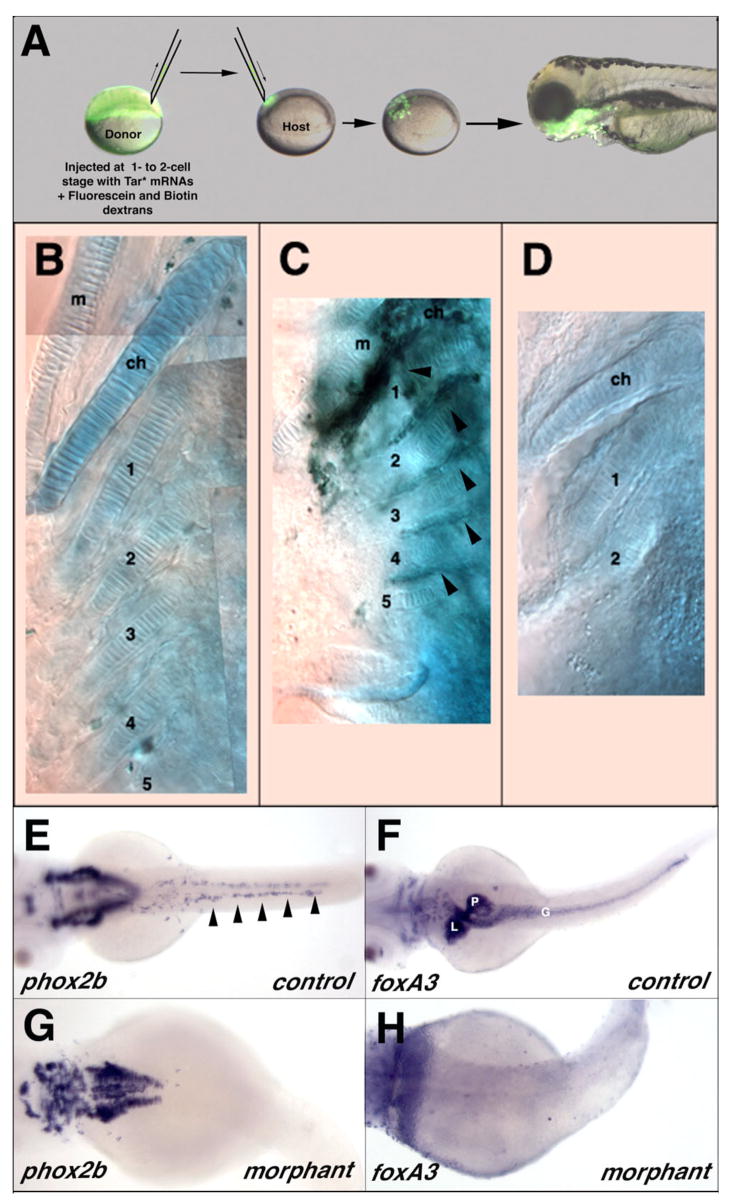
The zebrafish enteric nervous system (ENS), like those of all other vertebrate species, is principally derived from the vagal neural crest. The developmental controls that govern the specification and patterning of the ENS are not well understood. To identify genes required for the formation of the vertebrate ENS, we preformed a genetic screen in zebrafish. We isolated the lessen (lsn) mutation that has a significant reduction in the number of ENS neurons as well as defects in other cranial neural crest derived structures. We show that the lsn gene encodes a zebrafish orthologue of Trap100, one of the subunits of the TRAP/mediator transcriptional regulation complex. A point mutation in trap100 causes a premature stop codon that truncates the protein, causing a loss of function. Antisense-mediated knockdown of trap100 causes an identical phenotype to lsn. During development trap100 is expressed in a dynamic tissue-specific expression pattern consistent with its function in ENS and jaw cartilage development. Analysis of neural crest markers revealed that the initial specification and migration of the neural crest is unaffected in lsn mutants. Phosphohistone H3 immunocytochemistry revealed that there is a significant reduction in proliferation of ENS precursors in lsn mutants. Using cell transplantation studies, we demonstrate that lsn/trap100 acts cell autonomously in the pharyngeal mesendoderm and influences the development of neural crest derived cartilages secondarily. Furthermore, we show that endoderm is essential for ENS development. These studies demonstrate that lsn/trap100 is not required for initial steps of cranial neural crest development and migration, but is essential for later proliferation of ENS precursors in the intestine.
Figures
References
-
- Ajiro K, Yoda K, Utsumi K, Nishikawa Y. Alteration of cell cycle-dependent histone phosphorylations by okadaic acid. Induction of mitosis-specific H3 phosphorylation and chromatin condensation in mammalian interphase cells. J Biol Chem. 1996;271:13197–201. - PubMed
-
- Akoulitchev S, Chuikov S, Reinberg D. TFIIH is negatively regulated by cdk8-containing mediator complexes. Nature. 2000;407:102–6. - PubMed
-
- Alexander J, Rothenberg M, Henry GL, Stainier DY. casanova plays an early and essential role in endoderm formation in zebrafish. Dev Biol. 1999;215:343–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
