

Hyperdynamic plasticity of chromatin proteins in pluripotent embryonic stem cells
- PMID: 16399082
- PMCID: PMC1868458
- DOI: 10.1016/j.devcel.2005.10.017
Hyperdynamic plasticity of chromatin proteins in pluripotent embryonic stem cells
Erratum in
- Dev Cell. Jan 17;22(1):233-4
Abstract
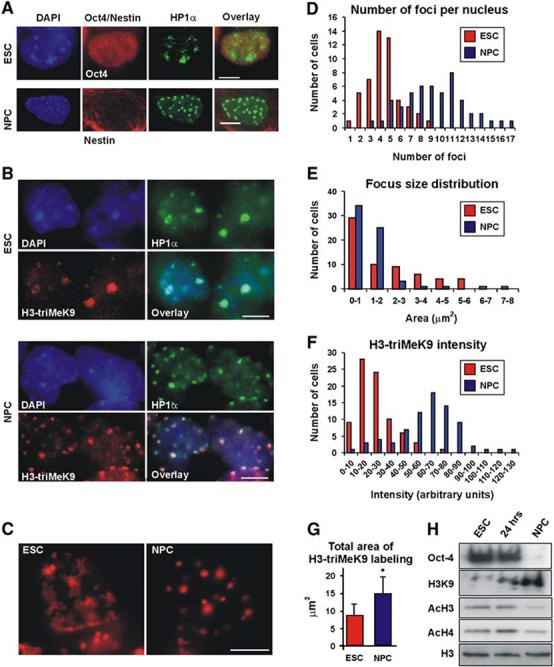
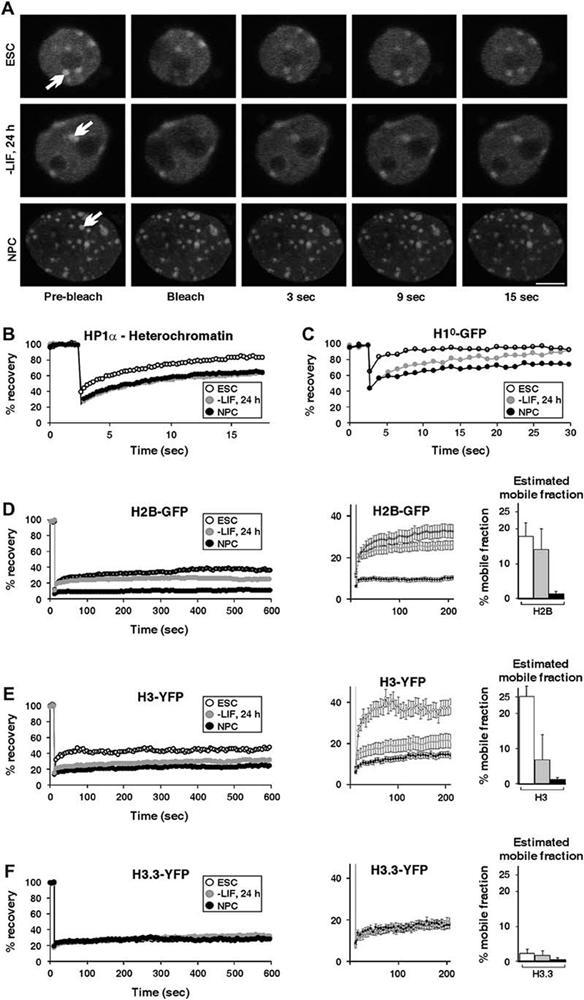
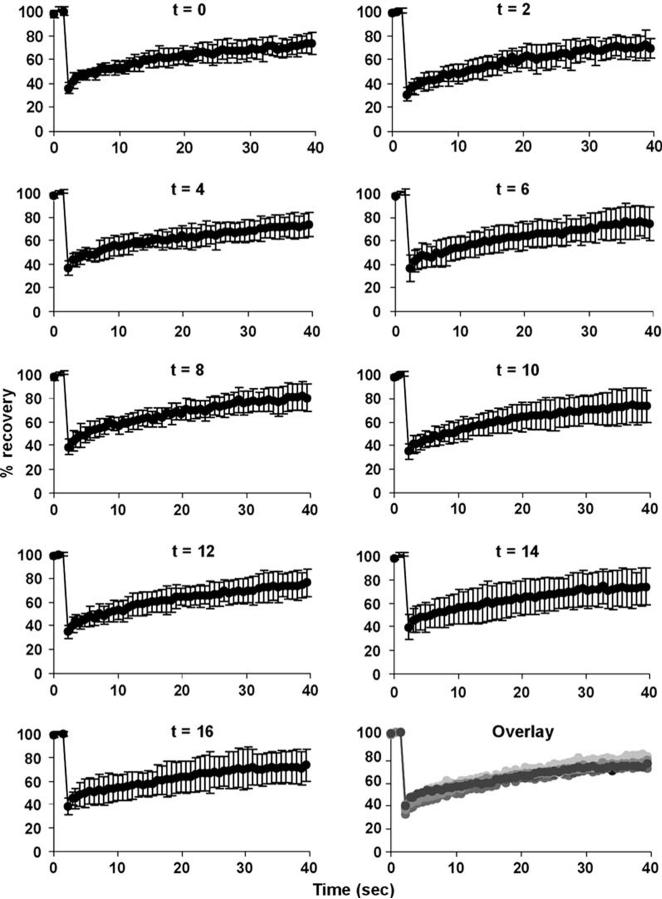
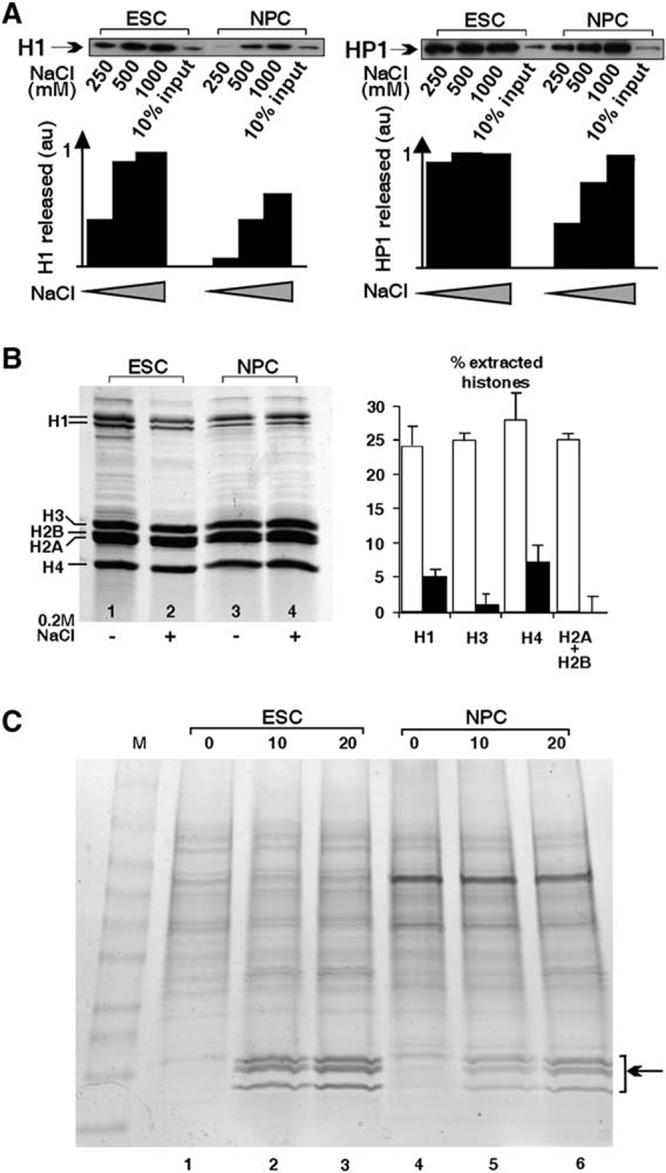
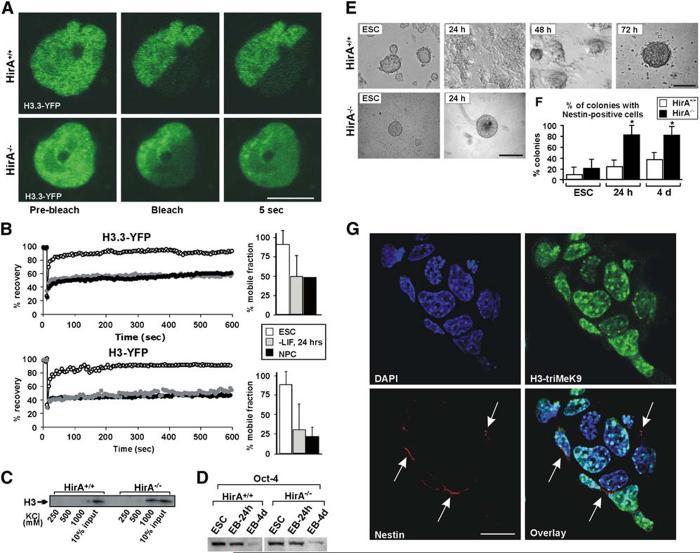
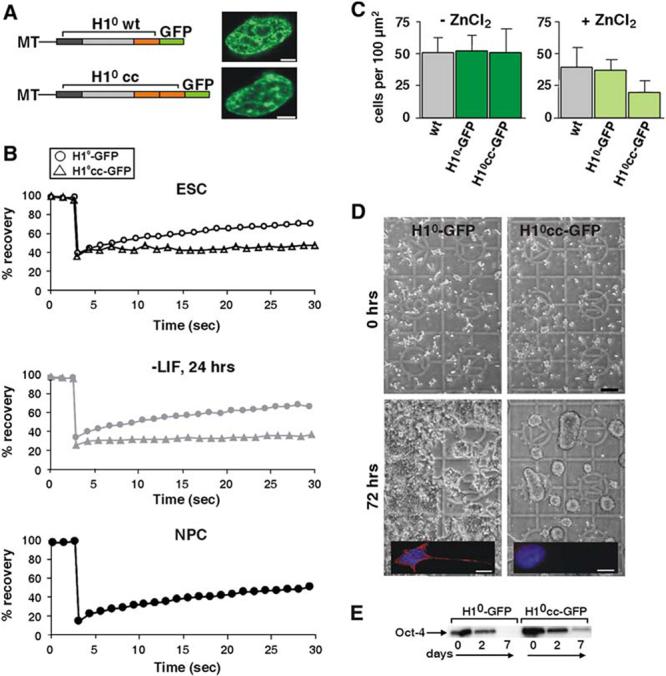
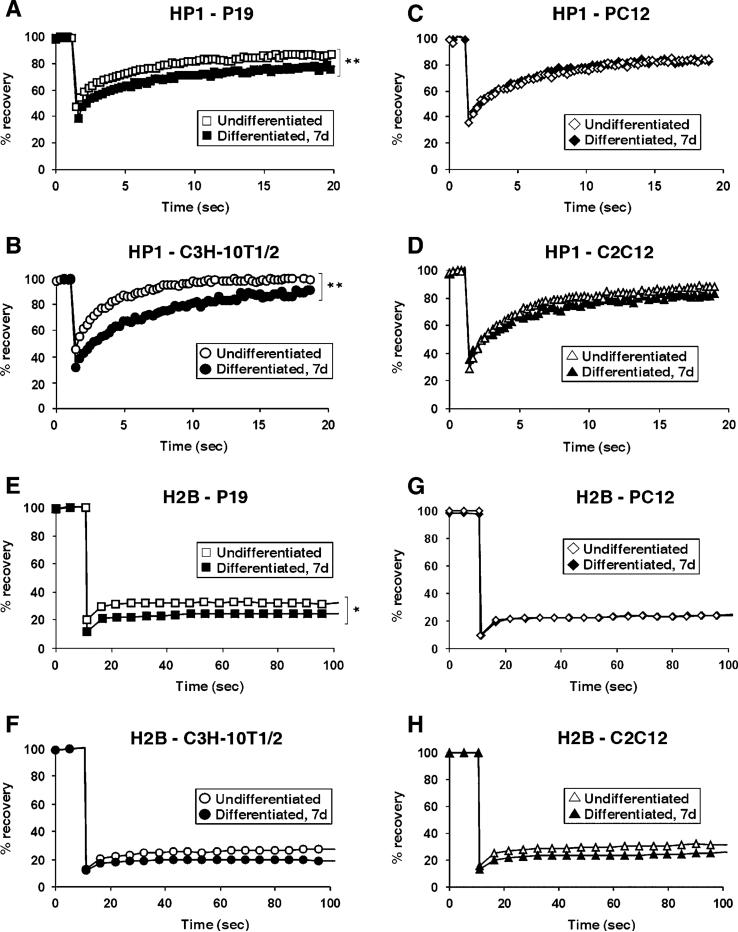
Differentiation of embryonic stem (ES) cells from a pluripotent to a committed state involves global changes in genome expression patterns. Gene activity is critically determined by chromatin structure and interactions of chromatin binding proteins. Here, we show that major architectural chromatin proteins are hyperdynamic and bind loosely to chromatin in ES cells. Upon differentiation, the hyperdynamic proteins become immobilized on chromatin. Hyperdynamic binding is a property of pluripotent cells, but not of undifferentiated cells that are already lineage committed. ES cells lacking the nucleosome assembly factor HirA exhibit elevated levels of unbound histones, and formation of embryoid bodies is accelerated. In contrast, ES cells, in which the dynamic exchange of H1 is restricted, display differentiation arrest. We suggest that hyperdynamic binding of structural chromatin proteins is a functionally important hallmark of pluripotent ES cells that contributes to the maintenance of plasticity in undifferentiated ES cells and to establishing higher-order chromatin structure.
Figures
Comment in
-
Breathing chromatin in pluripotent stem cells.Dev Cell. 2006 Jan;10(1):1-2. doi: 10.1016/j.devcel.2005.12.007. Dev Cell. 2006. PMID: 16399069
References
-
- Ahmad K, Henikoff S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell. 2002;9:1191–1200. - PubMed
-
- Ahn JI, Lee KH, Shin DM, Shim JW, Lee JS, Chang SY, Lee YS, Brownstein MJ, Lee SH. Comprehensive transcriptome analysis of differentiation of embryonic stem cells into midbrain and hindbrain neurons. Dev. Biol. 2004;265:491–501. - PubMed
-
- Cammas F, Oulad-Abdelghani M, Vonesch JL, Huss-Garcia Y, Chambon P, Losson R. Cell differentiation induces TIF1β association with centromeric heterochromatin via an HP1 interaction. J. Cell Sci. 2002;115:3439–3448. - PubMed
-
- Cheutin T, McNairn AJ, Jenuwein T, Gilbert DM, Singh PB, Misteli T. Maintenance of stable heterochromatin domains by dynamic HP1 binding. Science. 2003;299:721–725. - PubMed
-
- Eckfeldt CE, Mendenhall EM, Verfaillie CM. The molecular repertoire of the ‘almighty’ stem cell. Nat. Rev. Mol. Cell Biol. 2005;6:726–737. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
