

Parkinson's disease alpha-synuclein transgenic mice develop neuronal mitochondrial degeneration and cell death
- PMID: 16399671
- PMCID: PMC6381830
- DOI: 10.1523/JNEUROSCI.4308-05.2006
Parkinson's disease alpha-synuclein transgenic mice develop neuronal mitochondrial degeneration and cell death
Abstract
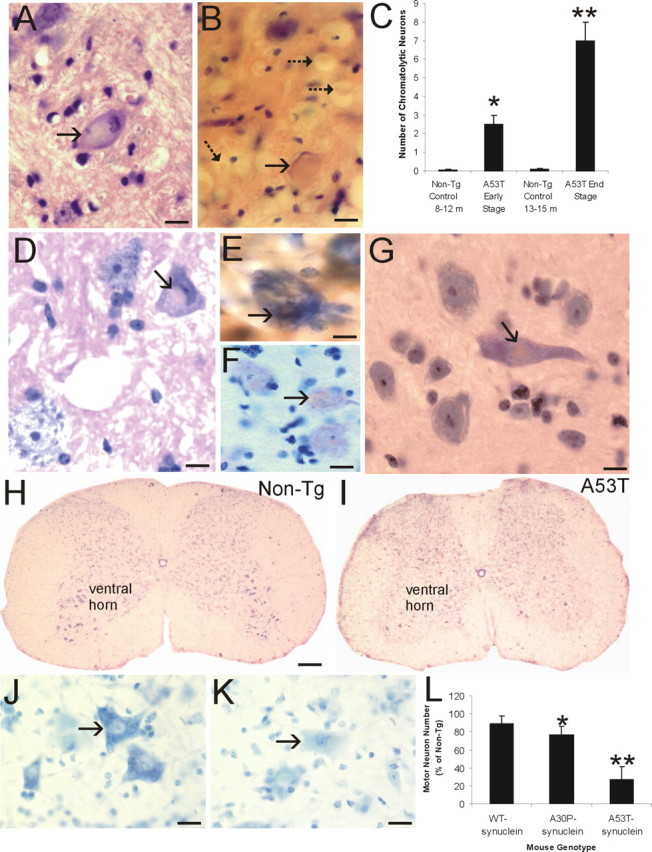
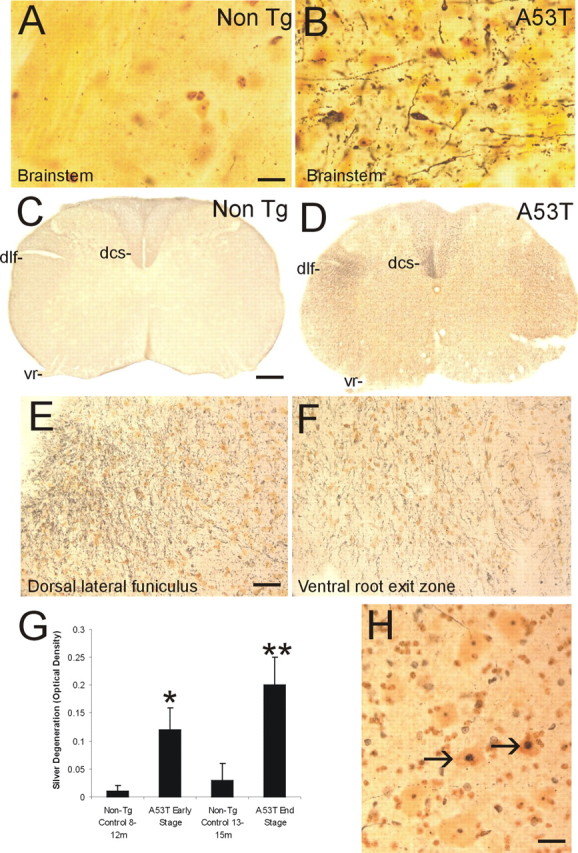
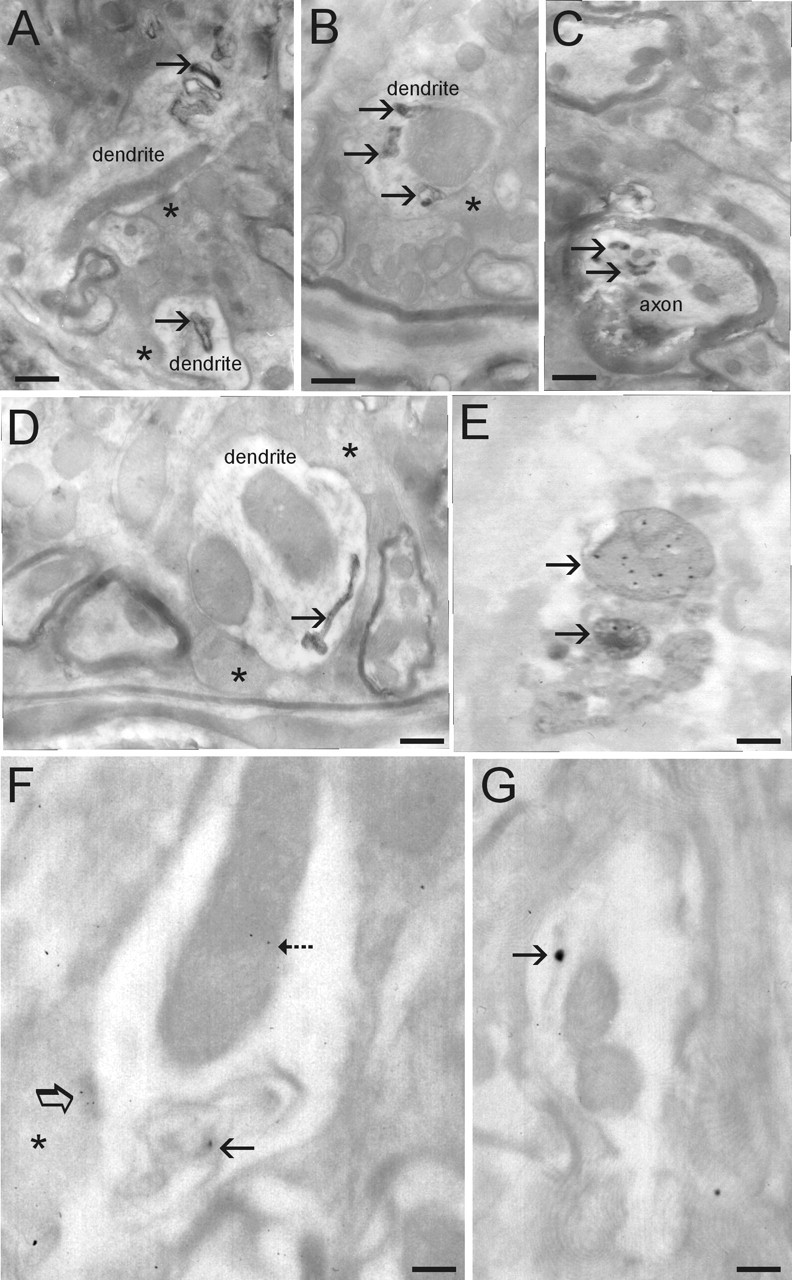
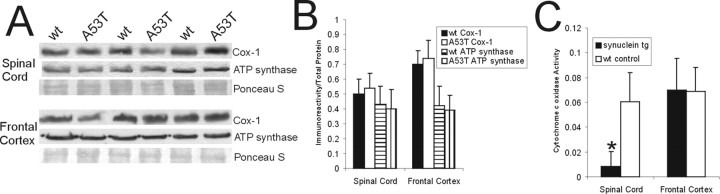
Alpha-synuclein (alpha-Syn) is enriched in nerve terminals. Two mutations in the alpha-Syn gene (Ala53--> Thr and Ala30--> Pro) occur in autosomal dominant familial Parkinson's disease. Mice overexpressing the human A53T mutant alpha-Syn develop a severe movement disorder, paralysis, and synucleinopathy, but the mechanisms are not understood. We examined whether transgenic mice expressing human wild-type or familial Parkinson's disease-linked A53T or A30P mutant alpha-syn develop neuronal degeneration and cell death. Mutant mice were examined at early- to mid-stage disease and at near end-stage disease. Age-matched nontransgenic littermates were controls. In A53T mice, neurons in brainstem and spinal cord exhibited large axonal swellings, somal chromatolytic changes, and nuclear condensation. Spheroid eosinophilic Lewy body-like inclusions were present in the cytoplasm of cortical neurons and spinal motor neurons. These inclusions contained human alpha-syn and nitrated synuclein. Motor neurons were depleted (approximately 75%) in A53T mice but were affected less in A30P mice. Axonal degeneration was present in many regions. Electron microscopy confirmed the cell and axonal degeneration and revealed cytoplasmic inclusions in dendrites and axons. Some inclusions were degenerating mitochondria and were positive for humanalpha-syn. Mitochondrial complex IV and V proteins were at control levels, but complex IV activity was reduced significantly in spinal cord. Subsets of neurons in neocortex, brainstem, and spinal cord ventral horn were positive for terminal deoxynucleotidyl transferase-mediated biotinylated UTP nick end labeling, cleaved caspase-3, and p53. Mitochondria in neurons had terminal deoxynucleotidyl transferase-mediated biotinylated UTP nick end labeling-positive matrices and p53 at the outer membrane. Thus, A53T mutant mice develop intraneuronal inclusions, mitochondrial DNA damage and degeneration, and apoptotic-like death of neocortical, brainstem, and motor neurons.
Figures

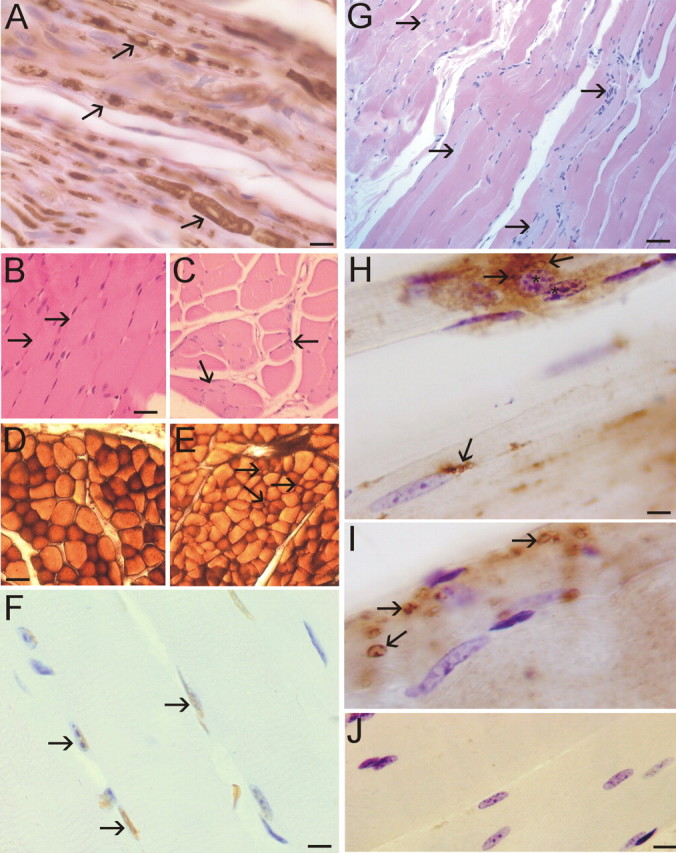
References
-
- Alves da Costa C, Paitel E, Vincent B, Checler F (2002) Alpha-synuclein lowers p53-dependent apoptotic of neuronal cell: abolishment by 6-hydroxydopamine and implication for Parkinson's disease. J Biol Chem 277: 50980–50984. - PubMed
-
- Braak H, Del Tredici K, Rüb U, de Vos RA, Jansen Steur ENH, Braak E (2003) Staging of brain pathology related to sporadic Parkinson's disease. Neurobiol Aging 24: 197–211. - PubMed
-
- Calhoun ME, Jucker M, Martin LJ, Thinakaran G, Price DL, Mouton PR (1996) Comparative evaluation of synaptophysin-based methods for quantification of synapses. J Neurocytol 25: 821–828. - PubMed
-
- Chayen J, Bitensky L, Butcher R (1973) Practical histochemistry. New York: Wiley.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous