

Sustained phosphorylation of Bid is a marker for resistance to Fas-induced apoptosis during chronic liver diseases
- PMID: 16401474
- PMCID: PMC1424224
- DOI: 10.1053/j.gastro.2005.10.012
Sustained phosphorylation of Bid is a marker for resistance to Fas-induced apoptosis during chronic liver diseases
Abstract
Background & aims: Increased rates of apoptosis have been reported to play a role in the pathophysiology of many disorders, including liver diseases. Conversely, genetic mutations that result in impairment of programmed cell death have been associated with cancer development. However, apoptosis resistance can also be the result of nongenetic stress adaptation, as seen in the cancer-prone metabolic liver disease hereditary tyrosinemia. To clarify whether stress-induced apoptosis resistance is a general feature of chronic liver diseases, an animal model of chronic cholestasis was examined.
Methods: Studies were performed with mice before and 2 weeks following bile duct ligation and with Fah-/- and Fah/p21-/- mice before and after NTBC withdrawal.
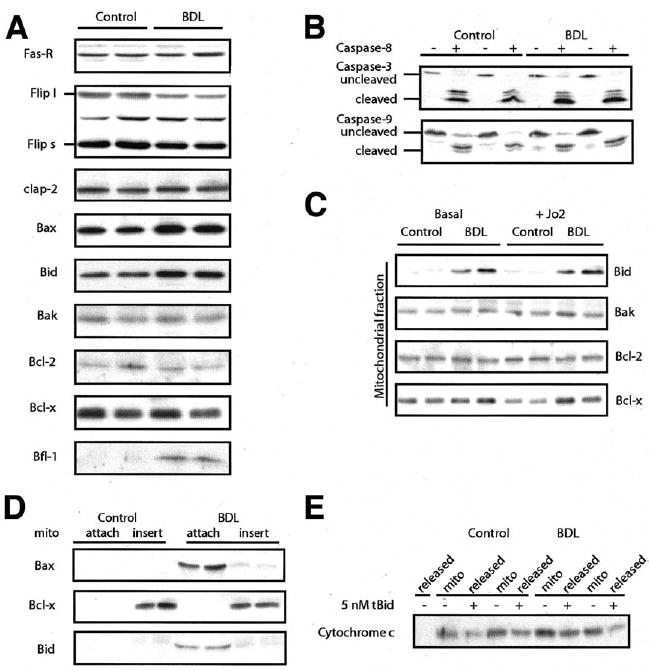
Results: Here we show that bile duct ligation induced profound resistance against Fas monoclonal antibody-mediated hepatocyte death. The apoptosis signaling pathway was blocked downstream of caspase-8 activation and proximal to mitochondrial cytochrome c release. In controls, activation of the Fas receptor resulted in rapid dephosphorylation of Bid and its subsequent cleavage, whereas Bid remained phosphorylated and uncleaved in chronic cholestasis and other models of hepatic apoptosis resistance.
Conclusions: We propose a model in which the phosphorylation status of Bid determines the apoptotic threshold of hepatocytes in vivo. Furthermore, resistance to apoptosis in chronic cholestasis may contribute to the long-term risk of cancer in this setting.
Figures




References
-
- Galle PR, Krammer PH. CD95-induced apoptosis in human liver disease. Semin Liver Dis. 1998;18:141–151. - PubMed
-
- Vogel A, van Den Berg IE, Al-Dhalimy M, Groopman J, Ou CN, Ryabinina O, Iordanov MS, Finegold M, Grompe M. Chronic liver disease in murine hereditary tyrosinemia type 1 induces resistance to cell death. Hepatology. 2004;39:433–443. - PubMed
-
- Findor J, He XS, Sord J, Terg R, Gershwin ME. Primary biliary cirrhosis and hepatocellular carcinoma. Autoimmun Rev. 2002;1:220–225. - PubMed
-
- Caballeria L, Pares A, Castells A, Gines A, Bru C, Rodes J. Hepatocellular carcinoma in primary biliary cirrhosis: similar incidence to that in hepatitis C virus-related cirrhosis. Am J Gastroenterol. 2001;96:1160–1163. - PubMed
-
- Keeffe EB, Pinson CW, Ragsdale J, Zonana J. Hepatocellular carcinoma in arteriohepatic dysplasia. Am J Gastroenterol. 1993;88:1446–1449. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
