

Receptor palmitoylation and ubiquitination regulate anthrax toxin endocytosis
- PMID: 16401723
- PMCID: PMC2063559
- DOI: 10.1083/jcb.200507067
Receptor palmitoylation and ubiquitination regulate anthrax toxin endocytosis
Abstract
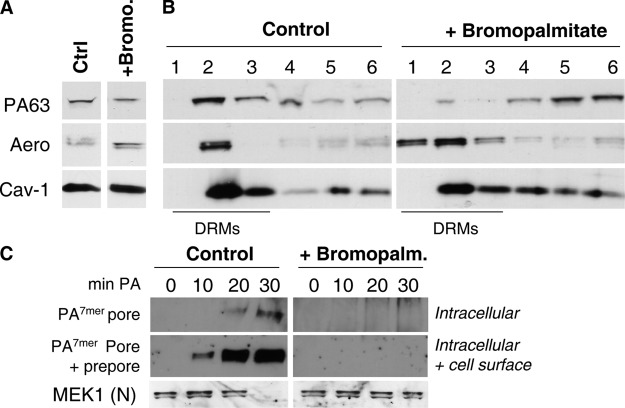
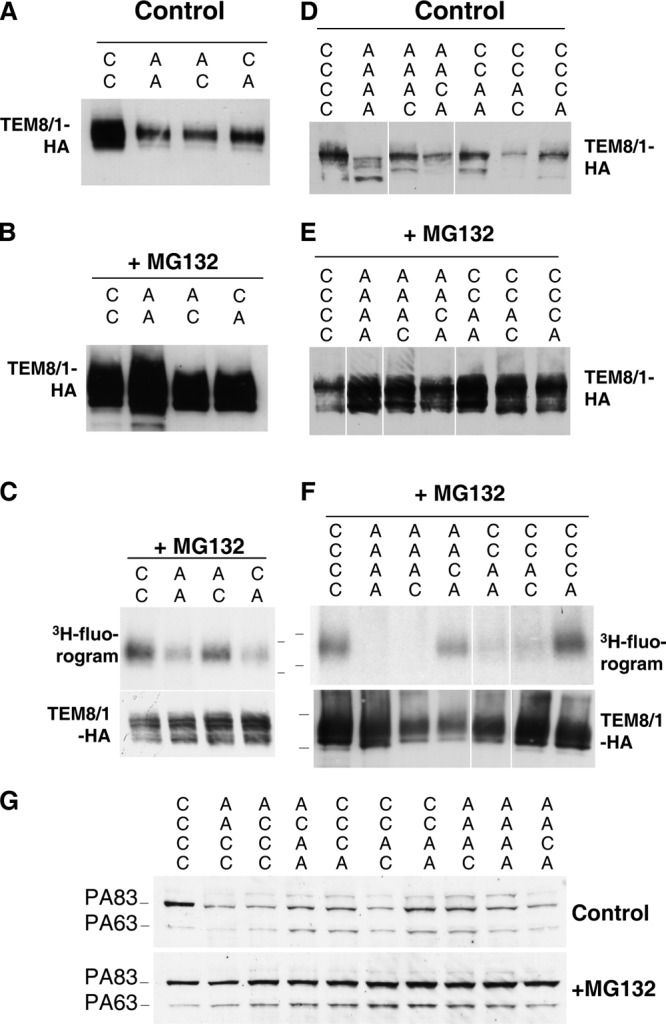
The anthrax toxin is composed of three independent polypeptide chains. Successful intoxication only occurs when heptamerization of the receptor-binding polypeptide, the protective antigen (PA), allows binding of the two enzymatic subunits before endocytosis. We show that this tailored behavior is caused by two counteracting posttranslational modifications in the cytoplasmic tail of PA receptors. The receptor is palmitoylated, and this unexpectedly prevents its association with lipid rafts and, thus, its premature ubiquitination. This second modification, which is mediated by the E3 ubiquitin ligase Cbl, only occurs in rafts and is required for rapid endocytosis of the receptor. As a consequence, cells expressing palmitoylation-defective mutant receptors are less sensitive to anthrax toxin because of a lower number of surface receptors as well as premature internalization of PA without a requirement for heptamerization.
Figures








References
-
- Abrami, L., N. Reig, and F.G. van der Goot. 2005. Anthrax toxin: the long and winding road that leads to the kill. Trends Microbiol. 13:72–78. - PubMed
-
- Alvarez, E., N. Girones, and R.J. Davis. 1990. Inhibition of the receptor-mediated endocytosis of diferric transferrin is associated with the covalent modification of the transferrin receptor with palmitic acid. J. Biol. Chem. 265:16644–16655. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
