

Purinergic signalling
- PMID: 16402102
- PMCID: PMC1760723
- DOI: 10.1038/sj.bjp.0706429
Purinergic signalling
Abstract
While there were early papers about the extracellular actions of purines, the role of ATP as a purinergic neurotransmitter in nonadrenergic, noncholinergic nerves in the gut and bladder in 1972 was a landmark discovery, although it met considerable resistance for the next 20 years. In the early 1990s, receptors for purines were cloned: four P1 receptor subtypes and seven P2X ionotropic and eight P2Y metabotropic receptor subtypes are currently recognized and characterized. The mechanisms underlying ATP release and breakdown are discussed. Purines and pyrimidines have major roles in the activities of non-neuronal cells as well as neurons. This includes fast signalling roles in exocrine and endocrine secretion, platelet aggregation, vascular endothelial cell-mediated vasodilation and nociceptive mechanosensory transduction, as well as acting as a cotransmitter and neuromodulator in most, if not all, nerve types in the peripheral and central nervous systems. More recently, slow (trophic) purinergic signalling has been implicated in cell proliferation, migration, differentiation and death in embryological development, wound healing, restenosis, atherosclerosis, ischaemia, cell turnover of epithelial cells in skin and visceral organs, inflammation, neuroprotection and cancer.
Figures
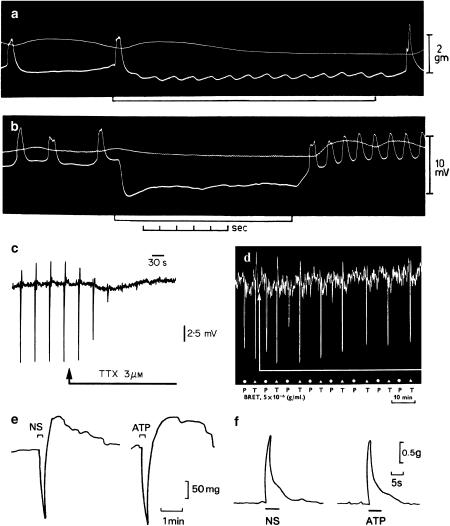
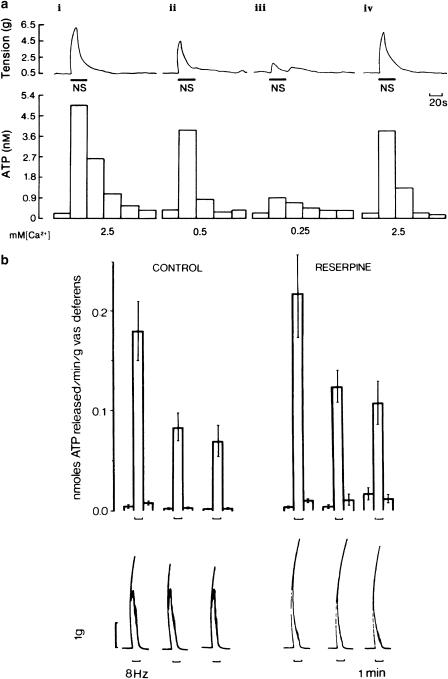
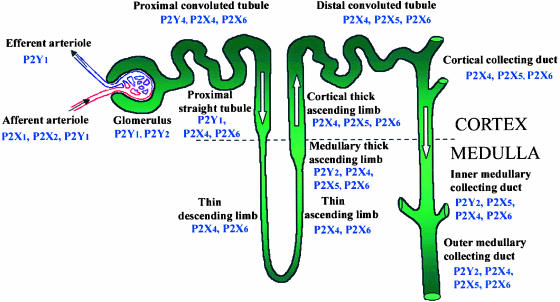
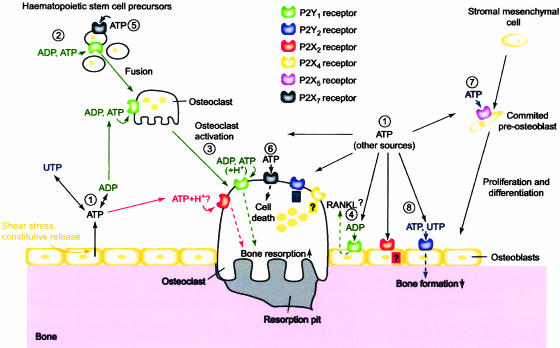
References
-
- ABBRACCHIO M.P., WILLIAMS M.(eds) (2001Handbook of Experimental Pharmacology: Purinergic and Pyrimidinergic Signalling, Vol. 151 (I and II) Berlin: Springer
-
- ABBRACCHIO M.P., BURNSTOCK G. Purinergic signalling: pathophysiological roles. Jpn. J. Pharmacol. 1998;78:113–145. - PubMed
-
- ABRAHAM E.H., SALIKHOVA A.Y., RAPAPORT E. ATP in the treatment of advanced cancer. Curr. Top. Membr. 2003;54:415–452.
-
- BAILEY M.A., TURNER C., HUS-CITHAREL A., MARCHETTI J., IMBERT-TEBOUL M., MILNER P., BURNSTOCK G., UNWIN R. P2Y receptors present in the native and isolated rat glomerulus. Nephron. Physiol. 2004;96:P79–P90. - PubMed
-
- BERNE R.M. Cardiac nucleotides in hypoxia: possible role in regulation of coronary blood flow. Am. J. Physiol. 1963;204:317–322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
