

Inefficient coupling between proton transport and ATP synthesis may be the pathogenic mechanism for NARP and Leigh syndrome resulting from the T8993G mutation in mtDNA
- PMID: 16402916
- PMCID: PMC1462703
- DOI: 10.1042/BJ20051748
Inefficient coupling between proton transport and ATP synthesis may be the pathogenic mechanism for NARP and Leigh syndrome resulting from the T8993G mutation in mtDNA
Abstract
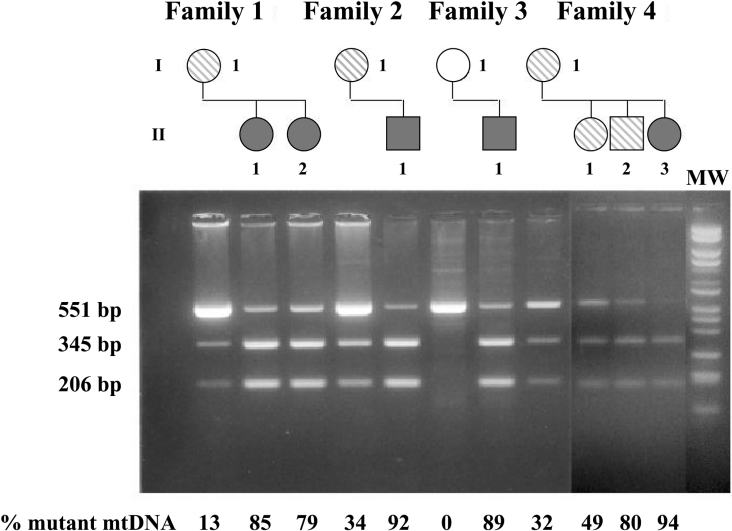
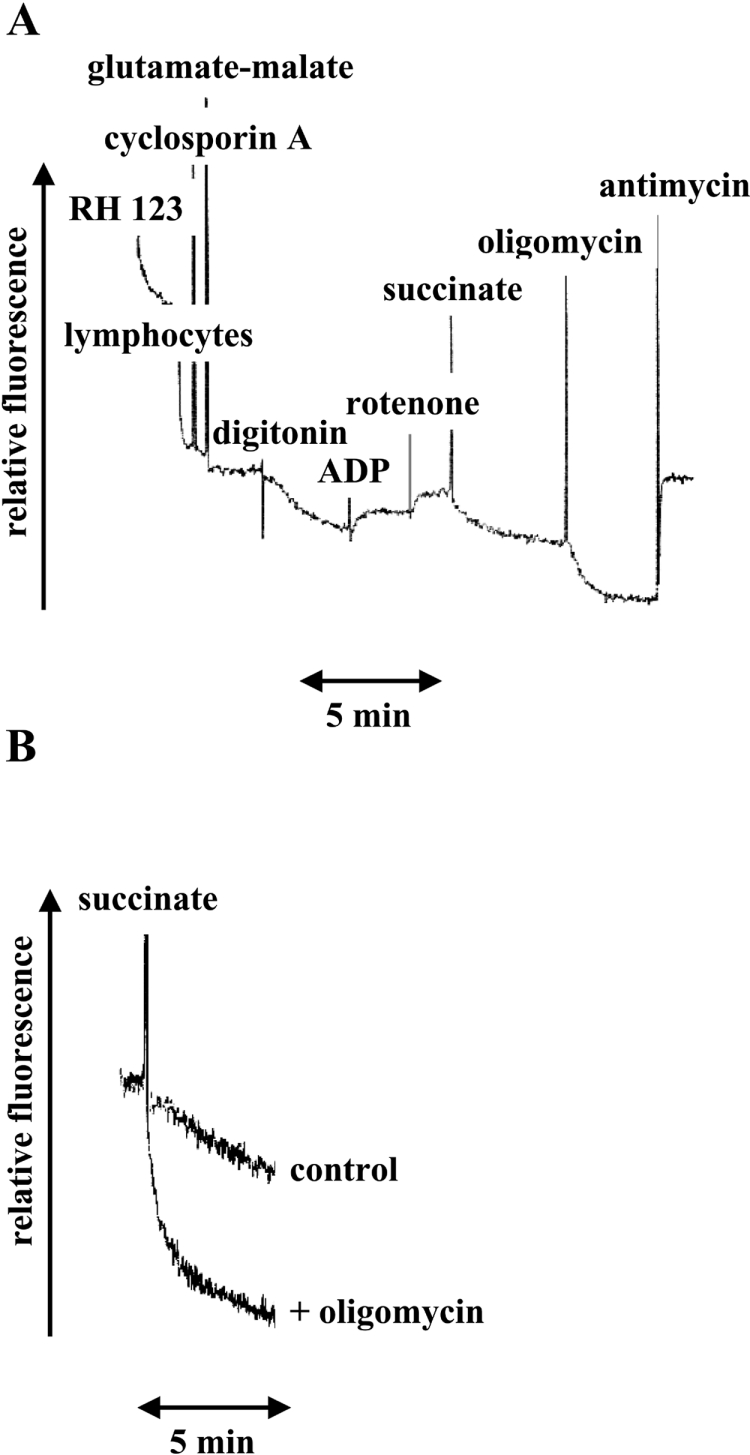
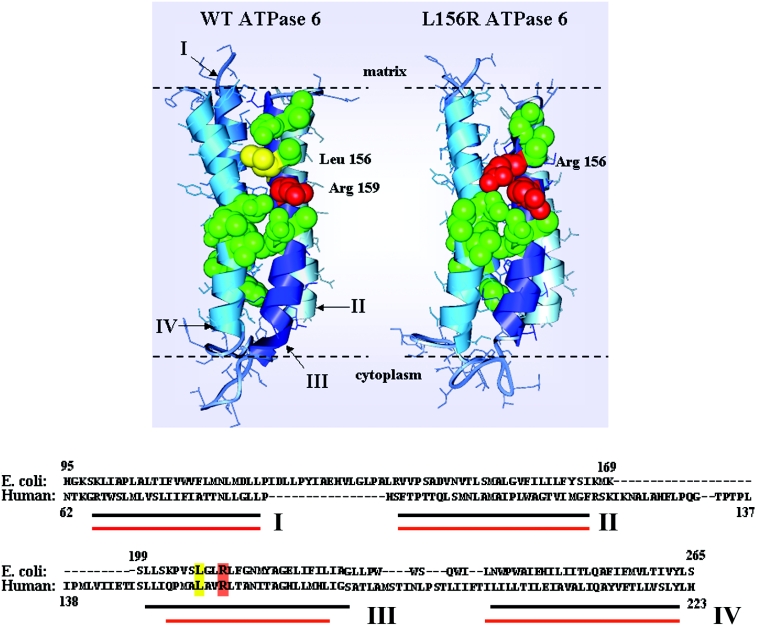
Mutations in the ATP6 gene of mtDNA (mitochondrial DNA) have been shown to cause several different neurological disorders. The product of this gene is ATPase 6, an essential component of the F1F0-ATPase. In the present study we show that the function of the F1F0-ATPase is impaired in lymphocytes from ten individuals harbouring the mtDNA T8993G point mutation associated with NARP (neuropathy, ataxia and retinitis pigmentosa) and Leigh syndrome. We show that the impaired function of both the ATP synthase and the proton transport activity of the enzyme correlates with the amount of the mtDNA that is mutated, ranging from 13-94%. The fluorescent dye RH-123 (Rhodamine-123) was used as a probe to determine whether or not passive proton flux (i.e. from the intermembrane space to the matrix) is affected by the mutation. Under state 3 respiratory conditions, a slight difference in RH-123 fluorescence quenching kinetics was observed between mutant and control mitochondria that suggests a marginally lower F0 proton flux capacity in cells from patients. Moreover, independent of the cellular mutant load the specific inhibitor oligomycin induced a marked enhancement of the RH-123 quenching rate, which is associated with a block in proton conductivity through F0 [Linnett and Beechey (1979) Inhibitors of the ATP synthethase system. Methods Enzymol. 55, 472-518]. Overall, the results rule out the previously proposed proton block as the basis of the pathogenicity of the mtDNA T8993G mutation. Since the ATP synthesis rate was decreased by 70% in NARP patients compared with controls, we suggest that the T8993G mutation affects the coupling between proton translocation through F0 and ATP synthesis on F1. We discuss our findings in view of the current knowledge regarding the rotary mechanism of catalysis of the enzyme.
Figures



References
-
- Boyer P. D. The ATP synthase – a splendid molecular machine. Annu. Rev. Biochem. 1997;66:717–749. - PubMed
-
- Weber J., Senior A. E. ATP synthesis driven by proton transport in F1F0-ATP synthase. FEBS Lett. 2003;545:61–70. - PubMed
-
- Collinson I. R., Skehel J. M., Fearnley I. M., Runswick M. J., Walker J. E. The F1Fo-ATPase complex from bovine heart mitochondria: the molar ratio of the subunits in the stalk region linking the F1 and F0 domains. Biochemistry. 1996;35:12640–12646. - PubMed
-
- Fillingame R. H., Angevine C. M., Dmitriev O. Y. Mechanics of coupling proton movements to c-ring rotation in ATP synthase. FEBS Lett. 2003;555:29–34. - PubMed
-
- Angevine C. M., Fillingame R. H. Aqueous access channels in subunit a of rotary ATP synthase. J. Biol. Chem. 2003;278:6066–6074. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical