

Alpha-gliadin genes from the A, B, and D genomes of wheat contain different sets of celiac disease epitopes
- PMID: 16403227
- PMCID: PMC1368968
- DOI: 10.1186/1471-2164-7-1
Alpha-gliadin genes from the A, B, and D genomes of wheat contain different sets of celiac disease epitopes
Abstract
Background: Bread wheat (Triticum aestivum) is an important staple food. However, wheat gluten proteins cause celiac disease (CD) in 0.5 to 1% of the general population. Among these proteins, the alpha-gliadins contain several peptides that are associated to the disease.
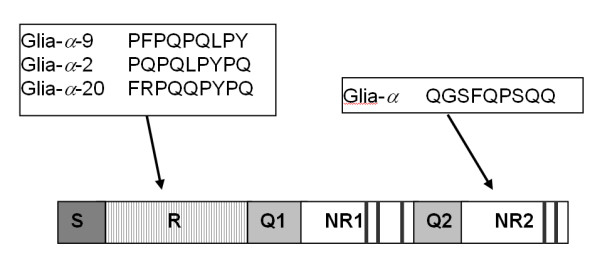
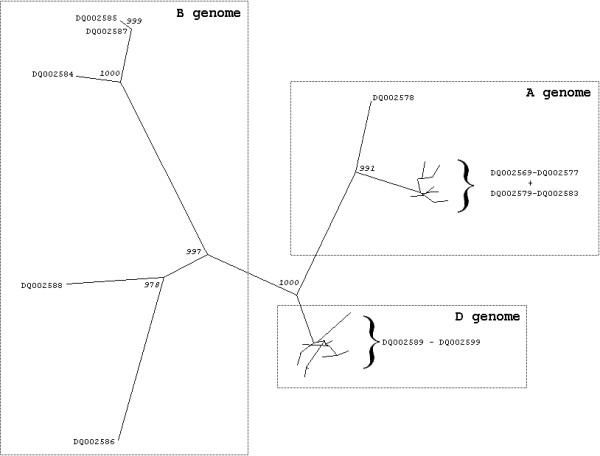
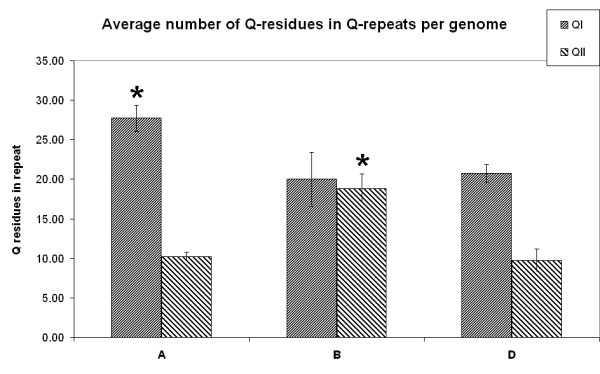
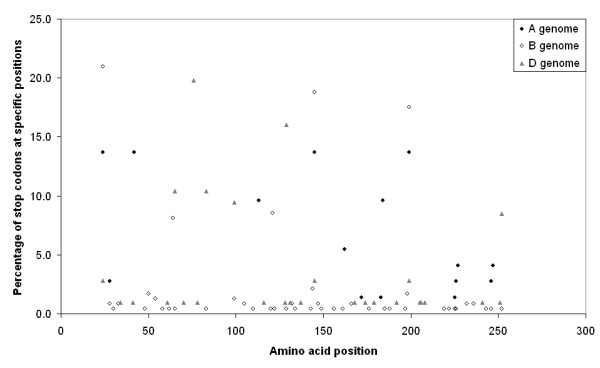
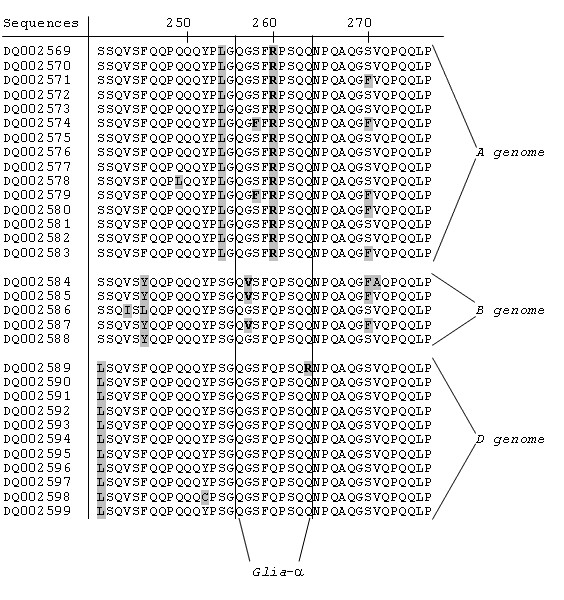
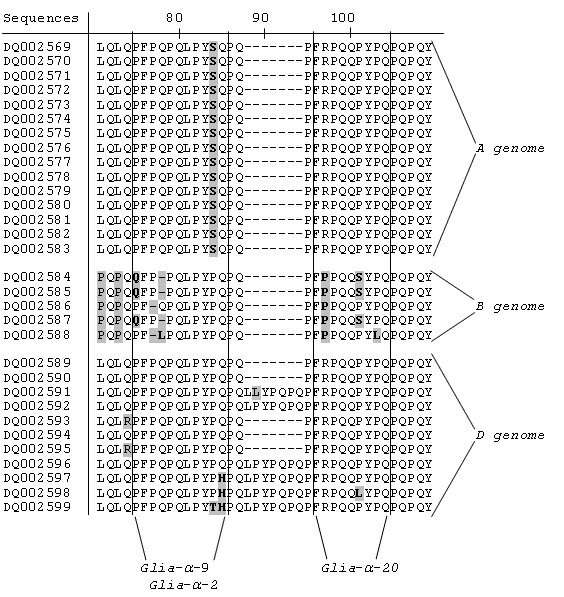
Results: We obtained 230 distinct alpha-gliadin gene sequences from severaldiploid wheat species representing the ancestral A, B, and D genomes of the hexaploid bread wheat. The large majority of these sequences (87%) contained an internal stop codon. All alpha-gliadin sequences could be distinguished according to the genome of origin on the basis of sequence similarity, of the average length of the polyglutamine repeats, and of the differences in the presence of four peptides that have been identified as T cell stimulatory epitopes in CD patients through binding to HLA-DQ2/8. By sequence similarity, alpha-gliadins from the public database of hexaploid T. aestivum could be assigned directly to chromosome 6A, 6B, or 6D. T. monococcum (A genome) sequences, as well as those from chromosome 6A of bread wheat, almost invariably contained epitope glia-alpha9 and glia-alpha20, but never the intact epitopes glia-alpha and glia-alpha2. A number of sequences from T. speltoides, as well as a number of sequences fromchromosome 6B of bread wheat, did not contain any of the four T cell epitopes screened for. The sequences from T. tauschii (D genome), as well as those from chromosome 6D of bread wheat, were found to contain all of these T cell epitopes in variable combinations per gene. The differences in epitope composition resulted mainly from point mutations. These substitutions appeared to be genome specific.
Conclusion: Our analysis shows that alpha-gliadin sequences from the three genomes of bread wheat form distinct groups. The four known T cell stimulatory epitopes are distributed non-randomly across the sequences, indicating that the three genomes contribute differently to epitope content. A systematic analysis of all known epitopes in gliadins and glutenins will lead to better understanding of the differences in toxicity among wheat varieties. On the basis of such insight, breeding strategies can be designed to generate less toxic varieties of wheat which may be tolerated by at least part of the CD patient population.
Figures
Similar articles
-
Molecular characterization of the celiac disease epitope domains in α-gliadin genes in Aegilops tauschii and hexaploid wheats (Triticum aestivum L.).Theor Appl Genet. 2010 Nov;121(7):1239-51. doi: 10.1007/s00122-010-1384-8. Epub 2010 Jun 17. Theor Appl Genet. 2010. PMID: 20556595
-
Diversification of the celiac disease α-gliadin complex in wheat: a 33-mer peptide with six overlapping epitopes, evolved following polyploidization.Plant J. 2015 Jun;82(5):794-805. doi: 10.1111/tpj.12851. Epub 2015 May 11. Plant J. 2015. PMID: 25864460
-
Celiac disease T-cell epitopes from gamma-gliadins: immunoreactivity depends on the genome of origin, transcript frequency, and flanking protein variation.BMC Genomics. 2012 Jun 22;13:277. doi: 10.1186/1471-2164-13-277. BMC Genomics. 2012. PMID: 22726570 Free PMC article.
-
Relation between gliadin structure and coeliac toxicity.Acta Paediatr Suppl. 1996 May;412:3-9. doi: 10.1111/j.1651-2227.1996.tb14239.x. Acta Paediatr Suppl. 1996. PMID: 8783747 Review.
-
Coeliac disease--a meeting point for genetics, immunology, and protein chemistry.Lancet. 2003 Apr 12;361(9365):1290-2. doi: 10.1016/s0140-6736(03)12989-3. Lancet. 2003. PMID: 12699968 Review.
Cited by
-
Cloning, expression, and evolutionary analysis of α-gliadin genes from Triticum and Aegilops genomes.J Appl Genet. 2013 May;54(2):157-67. doi: 10.1007/s13353-013-0139-z. Epub 2013 Mar 1. J Appl Genet. 2013. PMID: 23456845
-
Analytical and functional approaches to assess the immunogenicity of gluten proteins.Front Nutr. 2023 Jan 18;9:1049623. doi: 10.3389/fnut.2022.1049623. eCollection 2022. Front Nutr. 2023. PMID: 36741992 Free PMC article. Review.
-
Genome mapping of seed-borne allergens and immunoresponsive proteins in wheat.Sci Adv. 2018 Aug 17;4(8):eaar8602. doi: 10.1126/sciadv.aar8602. eCollection 2018 Aug. Sci Adv. 2018. PMID: 30128352 Free PMC article.
-
Effective shutdown in the expression of celiac disease-related wheat gliadin T-cell epitopes by RNA interference.Proc Natl Acad Sci U S A. 2010 Sep 28;107(39):17023-8. doi: 10.1073/pnas.1007773107. Epub 2010 Sep 9. Proc Natl Acad Sci U S A. 2010. PMID: 20829492 Free PMC article.
-
Celiac Immunogenic Potential of α-Gliadin Epitope Variants from Triticum and Aegilops Species.Nutrients. 2019 Jan 22;11(2):220. doi: 10.3390/nu11020220. Nutrients. 2019. PMID: 30678169 Free PMC article.
References
-
- Feldman M, Lupton FGH, Miller TE. In: Evolution of crop plant. Smartt J, Simmonds NW, editor. Harlow Essex: Longman Scientific & technical; 1995. pp. 184–192.
-
- von Buren M. Polymorphisms in two homeologous gamma-gliadin genes and the evolution of cultivated wheat. Genet Res Crop Evol. 2001;48:205–220. doi: 10.1023/A:1011213228222. - DOI
-
- Marino CL, Nelson JC, Lu YH, Sorrells ME, Leroy P, Tuleen NA, Lopes CR, Hart GE. Molecular genetic maps of the group 6 chromosomes of hexaploid wheat (Triticum aestivum L. em.Thell) Genome. 1996;39:359–366. - PubMed
-
- Harberd NP, Bartels D, Thompson RD. Analysis of the gliadin multigene locus in bread wheat using nullisomic-tetrasomic lines. Mol Gen Genet. 1985;198:234–242. doi: 10.1007/BF00383001. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
