

CaMKII and polo-like kinase 1 sequentially phosphorylate the cytostatic factor Emi2/XErp1 to trigger its destruction and meiotic exit
- PMID: 16407128
- PMCID: PMC1325965
- DOI: 10.1073/pnas.0509549102
CaMKII and polo-like kinase 1 sequentially phosphorylate the cytostatic factor Emi2/XErp1 to trigger its destruction and meiotic exit
Abstract
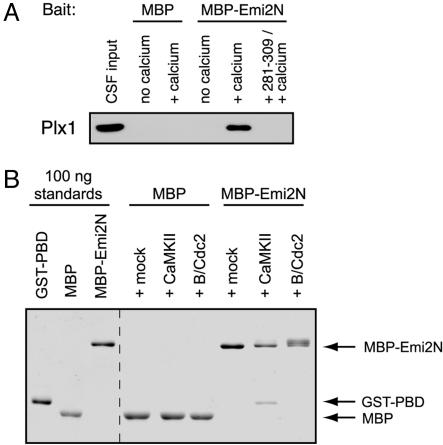
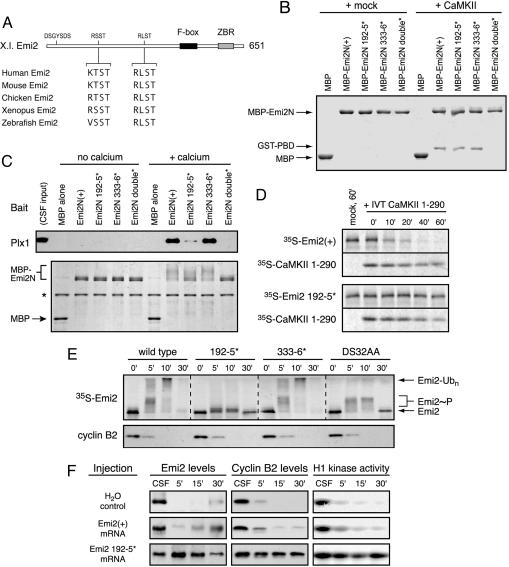
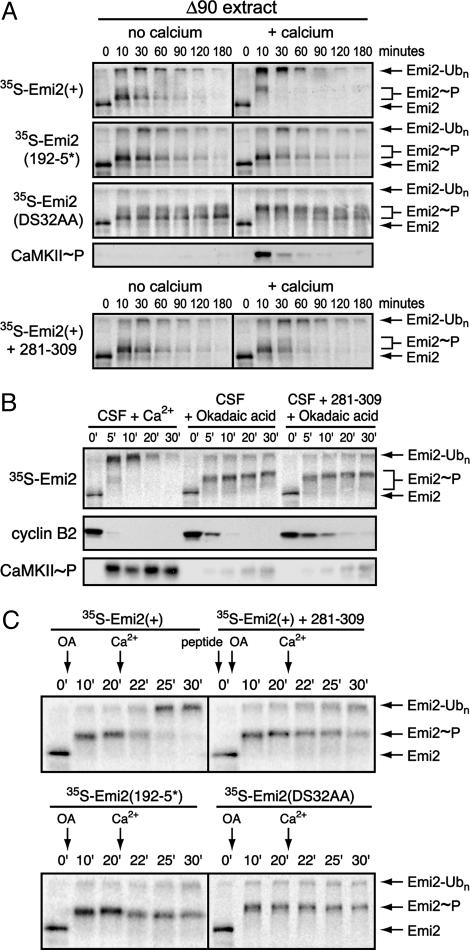
In vertebrate meiosis, unfertilized eggs are arrested in metaphase II by cytostatic factor (CSF), which is required to maintain mitotic cyclin-dependent kinase activity. Fertilization triggers a transient increase in cytosolic free Ca(2+), which leads to CSF inactivation and ubiquitin-dependent cyclin destruction through the anaphase promoting complex or cyclosome (APC/C). The Ca(2+)/calmodulin-dependent protein kinase II (CaMKII) and the Polo-like kinase Plx1 are essential factors for Ca(2+)-induced meiotic exit, but the critical targets of these kinases were unknown. The APC/C inhibitor Emi2 or XErp1 has recently been characterized as a pivotal CSF component, required to maintain metaphase II arrest and rapidly destroyed in response to Ca(2+) signaling through phosphorylation by Plx1 and ubiquitination by the SCF(betaTrCP) complex. An important question is how the increase in free Ca(2+) targets Plx1 activity toward Emi2. Here, we demonstrate that CaMKII is required for Ca(2+)-induced Emi2 destruction, and that CaMKII functions as a "priming kinase," directly phosphorylating Emi2 at a specific motif to induce a strong interaction with the Polo Box domain of Plx1. We show that the strict requirement for CaMKII to phosphorylate Emi2 is a specific feature of CSF arrest, and we also use phosphatase inhibitors to demonstrate an additional mode of Emi2 inactivation independent of its destruction. We firmly establish the CSF component Emi2 as the first-known critical and direct target of CaMKII in CSF release, providing a detailed molecular mechanism explaining how CaMKII and Plx1 coordinately direct APC/C activation and meiotic exit upon fertilization.
Figures

Similar articles
-
Calcium elevation at fertilization coordinates phosphorylation of XErp1/Emi2 by Plx1 and CaMK II to release metaphase arrest by cytostatic factor.Curr Biol. 2005 Aug 23;15(16):1458-68. doi: 10.1016/j.cub.2005.07.030. Curr Biol. 2005. PMID: 16040245
-
Calcium triggers exit from meiosis II by targeting the APC/C inhibitor XErp1 for degradation.Nature. 2005 Oct 13;437(7061):1048-52. doi: 10.1038/nature04093. Epub 2005 Aug 28. Nature. 2005. PMID: 16127448
-
A role for the anaphase-promoting complex inhibitor Emi2/XErp1, a homolog of early mitotic inhibitor 1, in cytostatic factor arrest of Xenopus eggs.Proc Natl Acad Sci U S A. 2005 Mar 22;102(12):4318-23. doi: 10.1073/pnas.0501108102. Epub 2005 Mar 7. Proc Natl Acad Sci U S A. 2005. PMID: 15753281 Free PMC article.
-
Across the meiotic divide - CSF activity in the post-Emi2/XErp1 era.J Cell Sci. 2008 Nov 1;121(Pt 21):3509-14. doi: 10.1242/jcs.036855. J Cell Sci. 2008. PMID: 18946022 Review.
-
[Molecular mechanisms for meiotic arrest that is cancelled by fertilization in vertebrate eggs].Tanpakushitsu Kakusan Koso. 2008 Mar;53(3):207-16. Tanpakushitsu Kakusan Koso. 2008. PMID: 18326293 Review. Japanese. No abstract available.
Cited by
-
Calcium and egg activation in Drosophila.Cell Calcium. 2013 Jan;53(1):10-5. doi: 10.1016/j.ceca.2012.11.008. Epub 2012 Dec 5. Cell Calcium. 2013. PMID: 23218670 Free PMC article. Review.
-
Regulation of store-operated Ca2+ entry during the cell cycle.J Cell Sci. 2010 Jul 1;123(Pt 13):2155-62. doi: 10.1242/jcs.069690. J Cell Sci. 2010. PMID: 20554894 Free PMC article.
-
Integrated multiple transcriptomes in oviductal tissue across the porcine estrous cycle reveal functional roles in oocyte maturation and transport.J Anim Sci. 2022 Feb 1;100(2):skab364. doi: 10.1093/jas/skab364. J Anim Sci. 2022. PMID: 34918099 Free PMC article.
-
Regulation of postsynaptic RapGAP SPAR by Polo-like kinase 2 and the SCFbeta-TRCP ubiquitin ligase in hippocampal neurons.J Biol Chem. 2008 Oct 24;283(43):29424-32. doi: 10.1074/jbc.M802475200. Epub 2008 Aug 22. J Biol Chem. 2008. PMID: 18723513 Free PMC article.
-
Identification of Cyclin-dependent Kinase 1 Specific Phosphorylation Sites by an In Vitro Kinase Assay.J Vis Exp. 2018 May 3;(135):57674. doi: 10.3791/57674. J Vis Exp. 2018. PMID: 29782014 Free PMC article.
References
-
- Runft, L. L., Jaffe, L. A. & Mehlmann, L. M. (2002) Dev. Biol. 245, 237–254. - PubMed
-
- Masui, Y. & Markert, C. L. (1971) J. Exp. Zool. 177, 129–145. - PubMed
-
- Sagata, N., Watanabe, N., Vande Woude, G. F. & Ikawa, Y. (1989) Nature 342, 512–518. - PubMed
-
- Tunquist, B. J. & Maller, J. L. (2003) Genes Dev. 17, 683–710. - PubMed
-
- Bhatt, R. R. & Ferrell, J. E., Jr. (1999) Science 286, 1362–1365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
