

High incidence of rapid telomere loss in telomerase-deficient Caenorhabditis elegans
- PMID: 16407328
- PMCID: PMC1326242
- DOI: 10.1093/nar/gkj417
High incidence of rapid telomere loss in telomerase-deficient Caenorhabditis elegans
Abstract
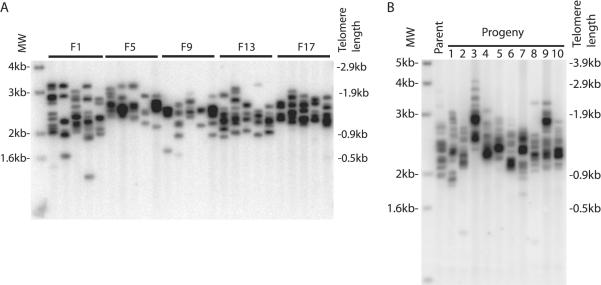
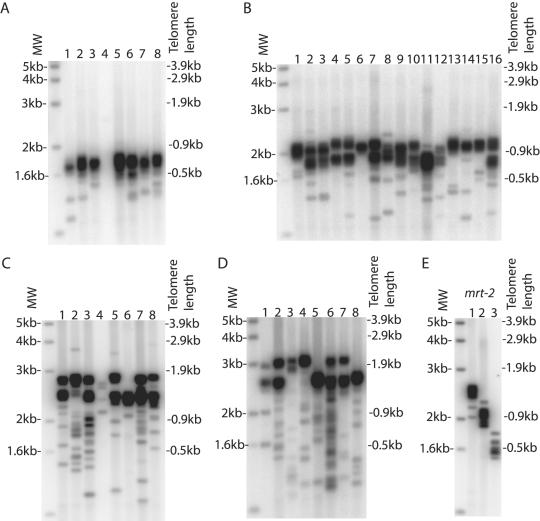
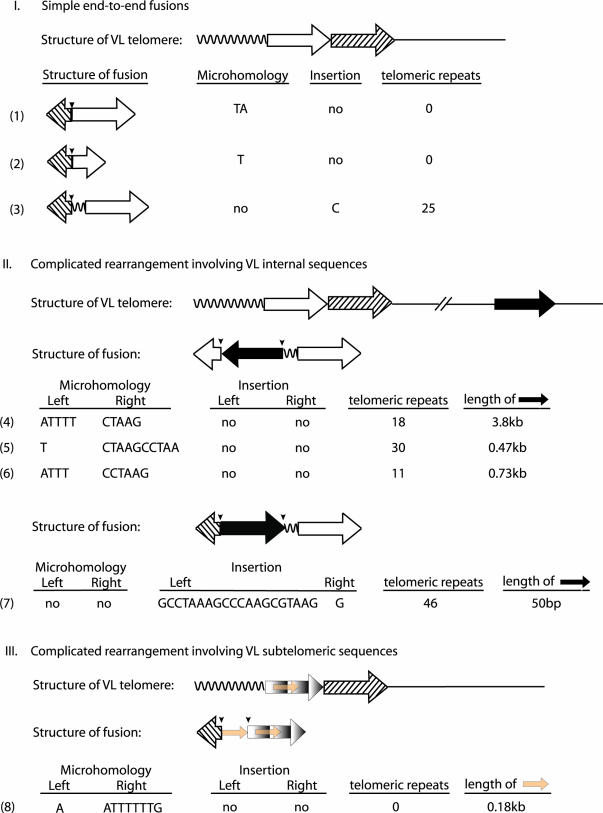
Telomerase is essential to maintain telomere length in most eukaryotes. Other functions for telomerase have been proposed but molecular mechanisms remain unclear. We studied Caenorhabditis elegans with a mutation in the trt-1 telomerase reverse transcriptase gene. Mutant animals showed a progressive decrease in brood size and typically failed to reproduce after five generations. Using PCR analysis to measure the length of individual telomere repeat tracks on the left arm of chromosome V we observed that trt-1 mutants lost approximately 125bp of telomeric DNA per generation. Chromosome fusions involving complex recombination reactions were observed in late generations. Strikingly, trt-1 mutant animals displayed a high frequency of telomeres with many fewer repeats than average. Such outlying short telomeres were not observed in mrt-2 mutants displaying progressive telomere loss very similar to trt-1 mutants. We speculate that, apart from maintaining the average telomere length, telomerase is required to prevent or repair sporadic telomere truncations that are unrelated to the typical 'end-replication' problems.
Figures

Similar articles
-
Strain-specific telomere length revealed by single telomere length analysis in Caenorhabditis elegans.Nucleic Acids Res. 2004 Jun 24;32(11):3383-91. doi: 10.1093/nar/gkh661. Print 2004. Nucleic Acids Res. 2004. PMID: 15247331 Free PMC article.
-
MRT-2 checkpoint protein is required for germline immortality and telomere replication in C. elegans.Nature. 2000 Jan 13;403(6766):159-64. doi: 10.1038/35003120. Nature. 2000. PMID: 10646593
-
trt-1 is the Caenorhabditis elegans catalytic subunit of telomerase.PLoS Genet. 2006 Feb;2(2):e18. doi: 10.1371/journal.pgen.0020018. Epub 2006 Feb 10. PLoS Genet. 2006. PMID: 16477310 Free PMC article.
-
Protection against chromosome degradation at the telomeres.Biochimie. 2008 Jan;90(1):41-59. doi: 10.1016/j.biochi.2007.07.008. Epub 2007 Jul 24. Biochimie. 2008. PMID: 17764802 Review.
-
Telomeres and aging.Physiol Rev. 2008 Apr;88(2):557-79. doi: 10.1152/physrev.00026.2007. Physiol Rev. 2008. PMID: 18391173 Review.
Cited by
-
Synthesis and antitumor mechanism of a new iron(iii) complex with 5,7-dichloro-2-methyl-8-quinolinol as ligands.Medchemcomm. 2017 Feb 1;8(3):633-639. doi: 10.1039/c6md00644b. eCollection 2017 Mar 1. Medchemcomm. 2017. PMID: 30108780 Free PMC article.
-
Maintenance of Genome Integrity by Mi2 Homologs CHD-3 and LET-418 in Caenorhabditis elegans.Genetics. 2018 Mar;208(3):991-1007. doi: 10.1534/genetics.118.300686. Epub 2018 Jan 16. Genetics. 2018. PMID: 29339410 Free PMC article.
-
Comparing effects of mTR and mTERT deletion on gene expression and DNA damage response: a critical examination of telomere length maintenance-independent roles of telomerase.Nucleic Acids Res. 2010 Jan;38(1):60-71. doi: 10.1093/nar/gkp855. Epub 2009 Oct 22. Nucleic Acids Res. 2010. PMID: 19850716 Free PMC article.
-
Extreme telomere length dimorphism in the Tasmanian devil and related marsupials suggests parental control of telomere length.PLoS One. 2012;7(9):e46195. doi: 10.1371/journal.pone.0046195. Epub 2012 Sep 25. PLoS One. 2012. PMID: 23049977 Free PMC article.
-
Telomere rapid deletion regulates telomere length in Arabidopsis thaliana.Mol Cell Biol. 2007 Mar;27(5):1706-15. doi: 10.1128/MCB.02059-06. Epub 2006 Dec 22. Mol Cell Biol. 2007. PMID: 17189431 Free PMC article.
References
-
- Blackburn E.H. Switching and signaling at the telomere. Cell. 2001;106:661–673. - PubMed
-
- d'Adda di Fagagna F., Reaper P.M., Clay-Farrace L., Fiegler H., Carr P., Von Zglinicki T., Saretzki G., Carter N.P., Jackson S.P. A DNA damage checkpoint response in telomere-initiated senescence. Nature. 2003;426:194–198. - PubMed
-
- Taggart A.K., Teng S.C., Zakian V.A. Est1p as a cell cycle-regulated activator of telomere-bound telomerase. Science. 2002;297:1023–1026. - PubMed
-
- Jacob N.K., Kirk K.E., Price C.M. Generation of telomeric G strand overhangs involves both G and C strand cleavage. Mol. Cell. 2003;11:1021–1032. - PubMed
-
- Masutomi K., Yu E.Y., Khurts S., Ben-Porath I., Currier J.L., Metz G.B., Brooks M.W., Kaneko S., Murakami S., DeCaprio J.A., et al. Telomerase maintains telomere structure in normal human cells. Cell. 2003;114:241–253. - PubMed
