

Erythroid progenitor renewal versus differentiation: genetic evidence for cell autonomous, essential functions of EpoR, Stat5 and the GR
- PMID: 16407844
- PMCID: PMC3035873
- DOI: 10.1038/sj.onc.1209308
Erythroid progenitor renewal versus differentiation: genetic evidence for cell autonomous, essential functions of EpoR, Stat5 and the GR
Abstract
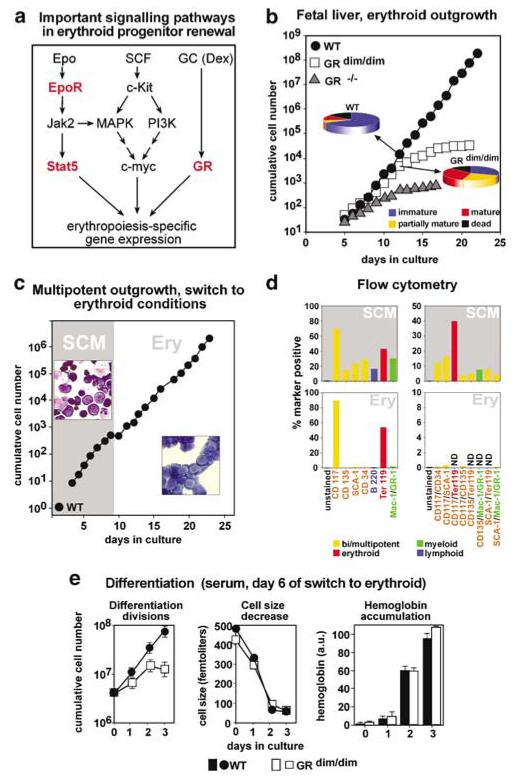
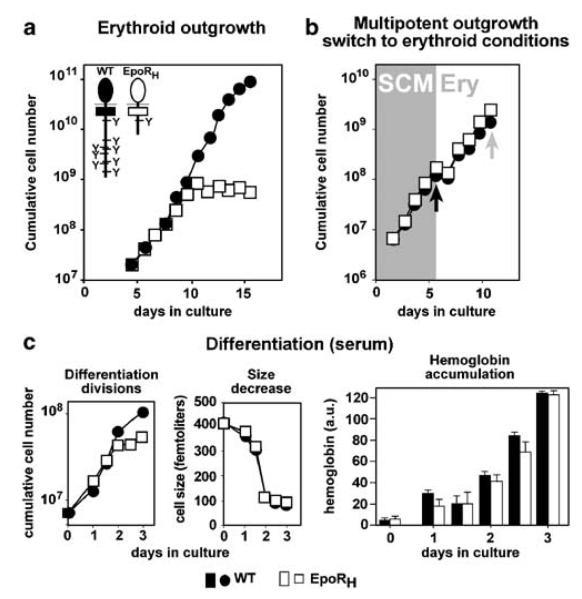
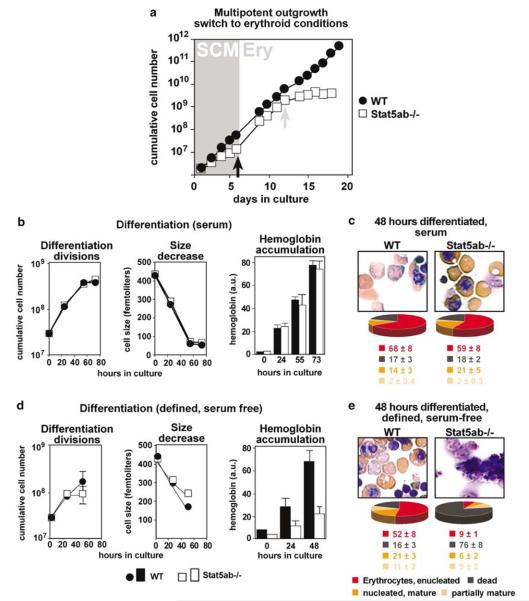
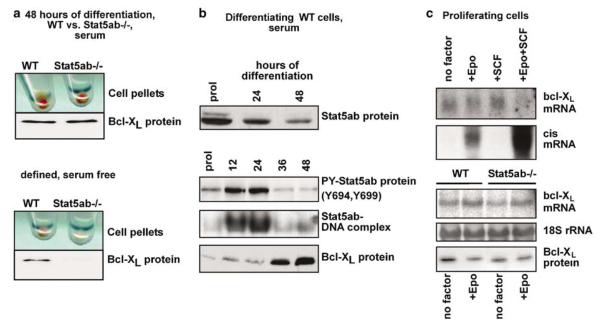
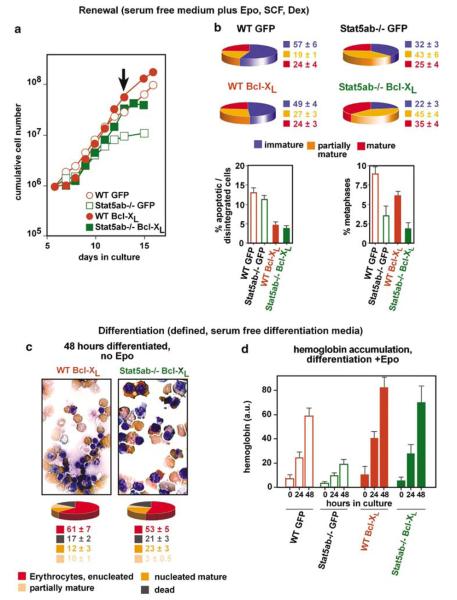
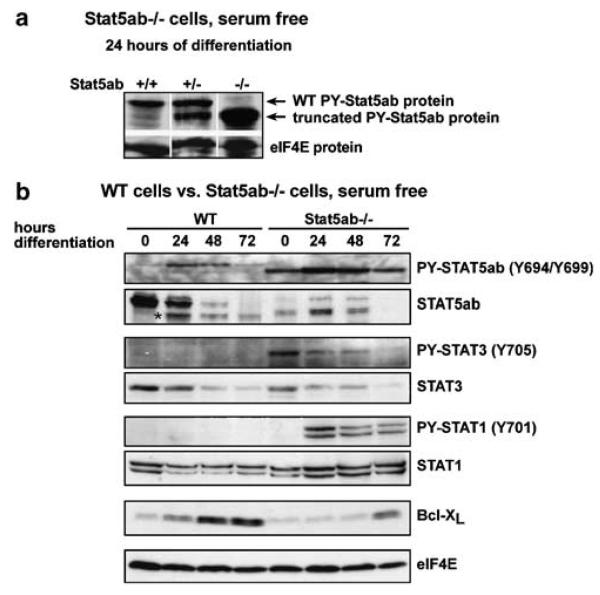
The balance between hematopoietic progenitor commitment and self-renewal versus differentiation is controlled by various transcriptional regulators cooperating with cytokine receptors. Disruption of this balance is increasingly recognized as important in the development of leukemia, by causing enhanced renewal and differentiation arrest. We studied regulation of renewal versus differentiation in primary murine erythroid progenitors that require cooperation of erythropoietin receptor (EpoR), the receptor tyrosine kinase c-Kit and a transcriptional regulator (glucocorticoid receptor; GR) for sustained renewal. However, mice defective for GR- (GR(dim/dim)), EpoR- (EpoR(H)) or STAT5ab function (Stat5ab(-/-)) show no severe erythropoiesis defects in vivo. Using primary erythroblast cultures from these mutants, we present genetic evidence that functional GR, EpoR, and Stat5 are essential for erythroblast renewal in vitro. Cells from GR(dim/dim), EpoR(H), and Stat5ab(-/-) mice showed enhanced differentiation instead of renewal, causing accumulation of mature cells and gradual proliferation arrest. Stat5ab was additionally required for Epo-induced terminal differentiation: differentiating Stat5ab(-/-) erythroblasts underwent apoptosis instead of erythrocyte maturation, due to absent induction of the antiapoptotic protein Bcl-X(L). This defect could be fully rescued by exogenous Bcl-X(L). These data suggest that signaling molecules driving leukemic proliferation may also be essential for prolonged self-renewal of normal erythroid progenitors.
Figures
References
-
- Bauer A, Gandrillon O, Samarut J, Beug H. In: Hematopoiesis: a developmental approach. Zon L, editor. Oxford University Press; Oxford: 2001. pp. 368–390.
-
- Beug H, Bauer A, Dolznig H, von Lindern M, Lobmayer L, Mellitzer G, et al. Biochim Biophys Acta. 1996;1288:M35–M47. - PubMed
-
- Beug H, Dahl R, Steinlein P, Meyer S, Deiner E, Hayman MH. Oncogene. 1995;11:59–72. - PubMed
-
- Blau HM, Brazelton TR, Weimann JM. Cell. 2001;105:829–841. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
