

SMUG1 is able to excise uracil from immunoglobulin genes: insight into mutation versus repair
- PMID: 16407970
- PMCID: PMC1383525
- DOI: 10.1038/sj.emboj.7600939
SMUG1 is able to excise uracil from immunoglobulin genes: insight into mutation versus repair
Abstract
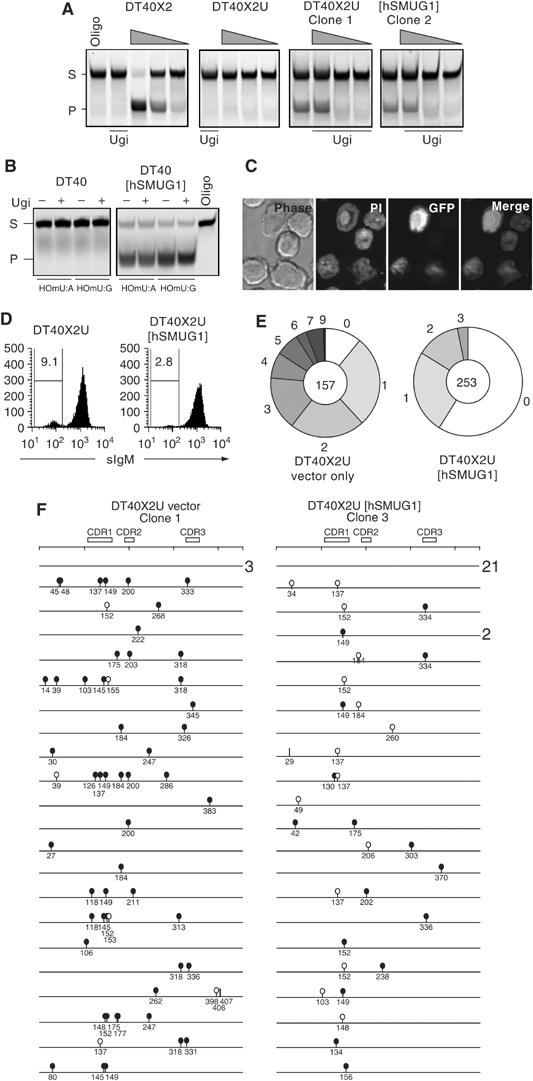
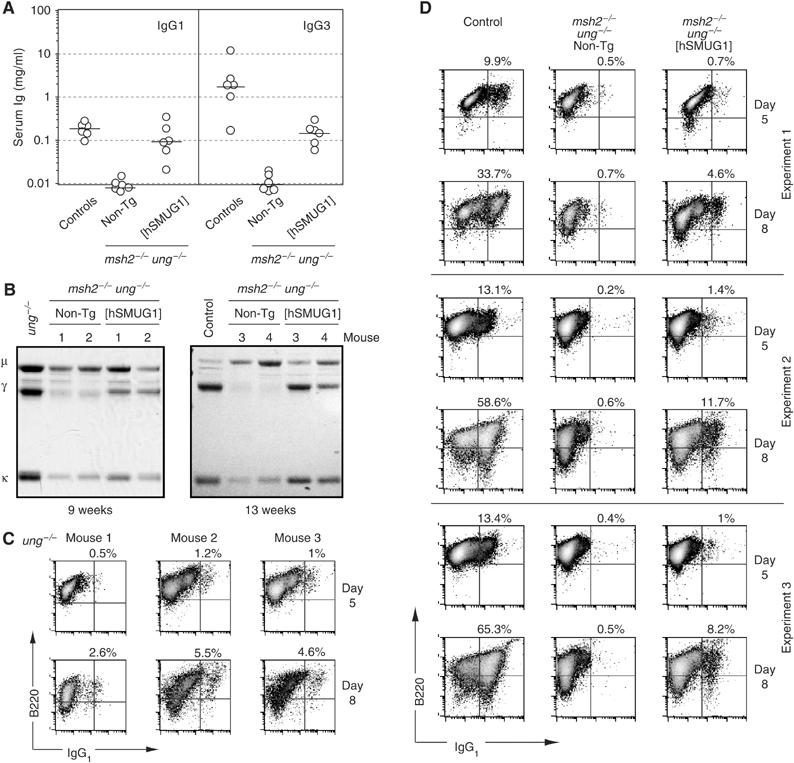
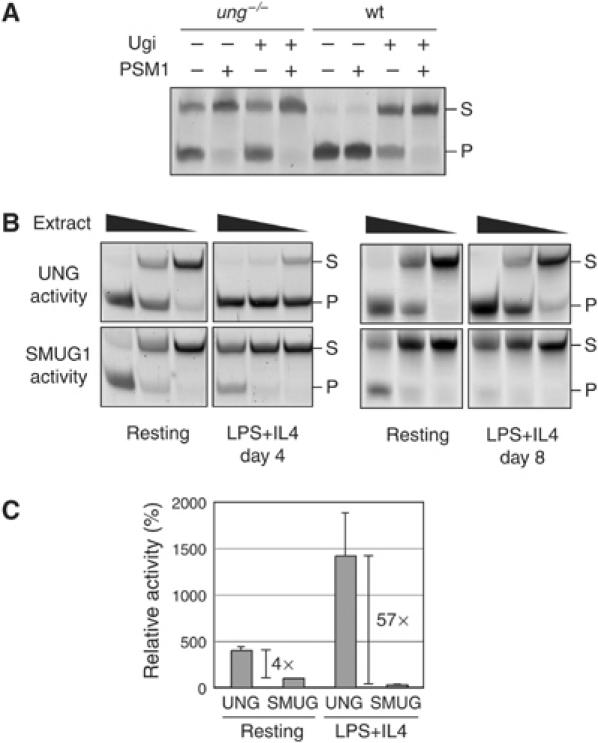
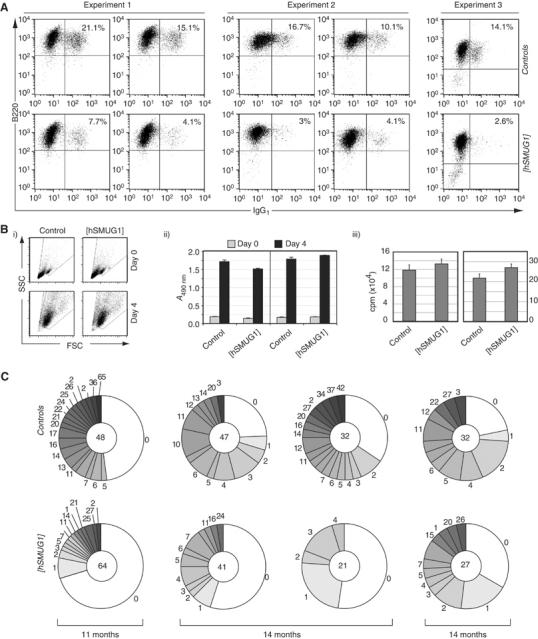
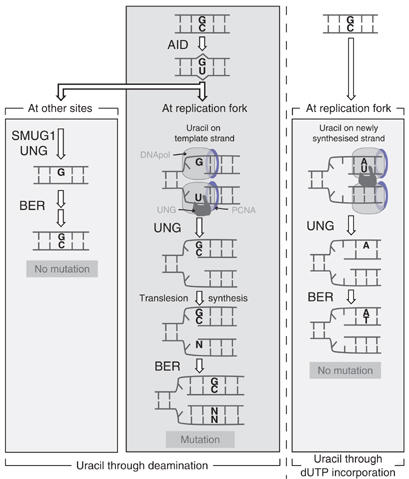
Mammals harbour multiple enzymes capable of excising uracil from DNA, although their distinct physiological roles remain uncertain. One of them (UNG) plays a critical role in antibody gene diversification, as UNG deficiency alone is sufficient to perturb the process. Here, we show this unique requirement for UNG does not reflect the fact that other glycosylases are unable to access the U:G lesion. SMUG1, if overexpressed, can partially substitute for UNG to assist antibody diversification as judged by its effect on somatic hypermutation patterns (in both DT40 B cells and mice) as well as a restoration of isotype switching in SMUG-transgenic msh2-/- ung-/- mice. However, SMUG1 plays little natural role in antibody diversification because (i) it is diminishingly expressed during B-cell activation and (ii) even if overexpressed, SMUG1 more appears to favour conventional repair of the uracil lesion than assist diversification. The distinction between UNG and overexpressed SMUG1 regarding the balance between antibody diversification and non-mutagenic repair of the U:G lesion could reflect the association of UNG (but not SMUG1) with sites of DNA replication.
Figures
References
-
- Akbari M, Otterlei M, Pena-Diaz J, Aas PA, Kavli B, Liabakk NB, Hagen L, Imai K, Durandy A, Slupphaug G, Krokan HE (2004) Repair of U/G and U/A in DNA by UNG2-associated repair complexes takes place predominantly by short-patch repair both in proliferating and growth-arrested cells. Nucleic Acids Res 32: 5486–5498 - PMC - PubMed
-
- Arakawa H, Hauschild J, Buerstedde J M (2002) Requirement of the activation-induced deaminase (AID) gene for immunoglobulin gene conversion. Science 295: 1301–1306 - PubMed
-
- Begum NA, Kinoshita K, Kakazu N, Muramatsu M, Nagaoka H, Shinkura R, Biniszkiewicz D, Boyer LA, Jaenisch R, Honjo T (2004) Uracil DNA glycosylaser activity is dispensable for immunoglobulin class switch. Science 305: 1160–1163 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
