

Infectious molecular clones from a simian immunodeficiency virus-infected rapid-progressor (RP) macaque: evidence of differential selection of RP-specific envelope mutations in vitro and in vivo
- PMID: 16415023
- PMCID: PMC1346964
- DOI: 10.1128/JVI.80.3.1463-1475.2006
Infectious molecular clones from a simian immunodeficiency virus-infected rapid-progressor (RP) macaque: evidence of differential selection of RP-specific envelope mutations in vitro and in vivo
Abstract
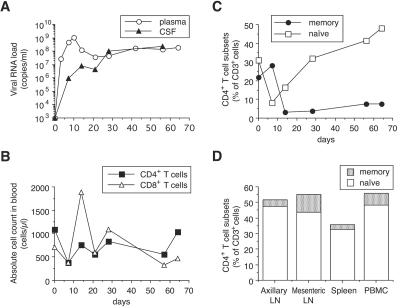
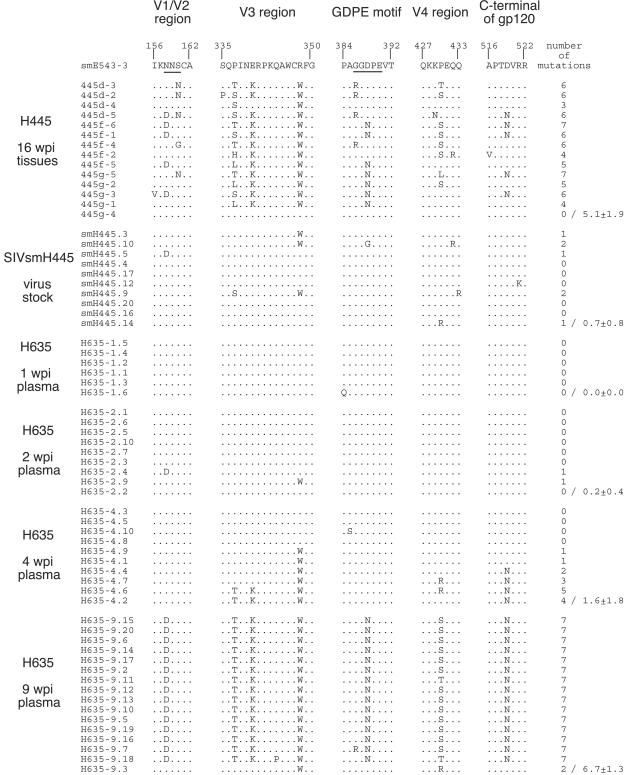
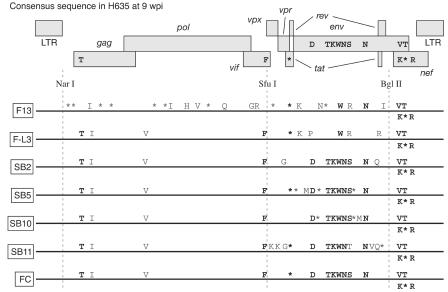
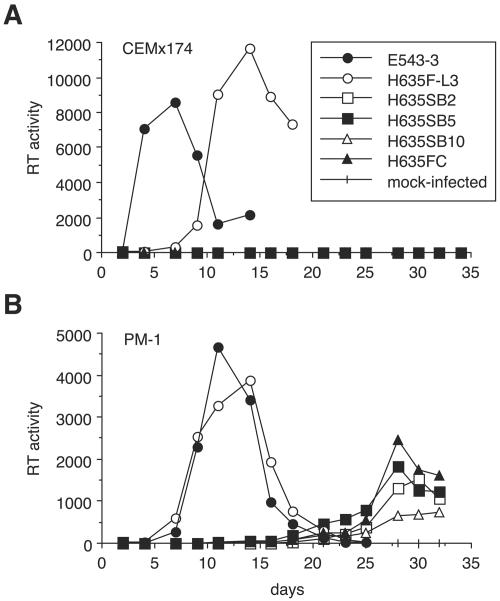
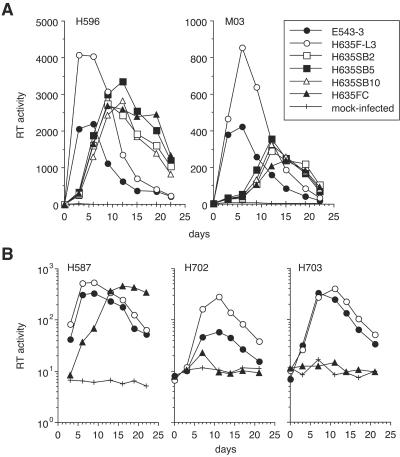
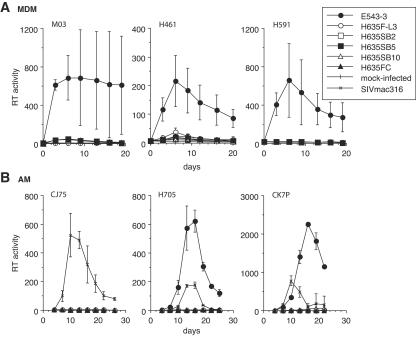
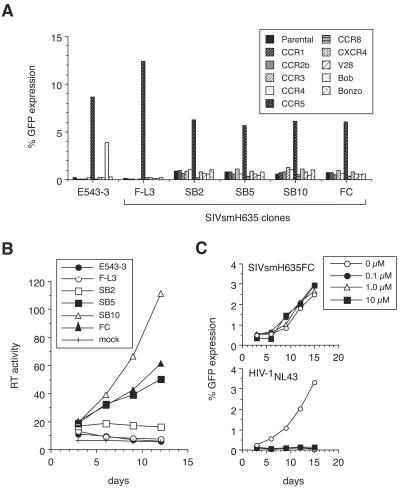
A minor fraction of simian immunodeficiency virus (SIV)-infected macaques progress rapidly to AIDS in the absence of SIV-specific immune responses. Common mutations in conserved residues of env in three SIVsmE543-3-infected rapid-progressor (RP) macaques suggest the evolution of a common viral variant in RP macaques. The goal of the present study was to analyze the biological properties of these variants in vitro and in vivo through the derivation of infectious molecular clones. Virus isolated from a SIVsmE543-3-infected RP macaque, H445 was used to inoculate six naive rhesus macaques. Although RP-specific mutations dominated in H445 tissues, they represented only 10% of the population of the virus stock, suggesting a selective disadvantage in vitro. Only one of these macaques (H635) progressed rapidly to AIDS. Plasma virus during primary infection of H635 was similar to the inoculum. However, RP-specific mutations were apparently rapidly reselected by 4 to 9 weeks postinfection. Terminal plasma from H635 was used as a source of viral RNA to generate seven full-length, infectious molecular clones. With the exception of one clone, which was similar to SIVsmE543-3, clones with RP-specific mutations replicated with delayed kinetics in rhesus peripheral blood mononuclear cells and human T-cell lines. None of the clones replicated in monocyte-derived or alveolar macrophages, and all used CCR5 as their major coreceptor. RP variants appear to be well adapted to replicate in vivo in RP macaques but are at a disadvantage in tissue culture compared to their parent, SIVsmE543-3. Therefore, tissue culture may not provide a good surrogate for replication of RP variants in macaques. These infectious clones will provide a valuable reagent to study the roles of specific viral variants in rapid progression in vivo.
Figures
References
-
- Anderson, M. G., D. Hauer, D. P. Sharma, S. V. Joag, O. Narayan, M. C. Zink, and J. E. Clements. 1993. Analysis of envelope changes acquired by SIVmac239 during neuroadaption in rhesus macaques. Virology 195:616-626. - PubMed
-
- Baskin, G. B., M. Murphey-Corb, E. A. Watson, and L. N. Martin. 1988. Necropsy findings in rhesus monkeys experimentally infected with cultured simian immunodeficiency virus (SIV)/delta. Vet. Pathol. 25:456-467. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
