

Mutational analysis of the herpes simplex virus triplex protein VP19C
- PMID: 16415029
- PMCID: PMC1346951
- DOI: 10.1128/JVI.80.3.1537-1548.2006
Mutational analysis of the herpes simplex virus triplex protein VP19C
Abstract
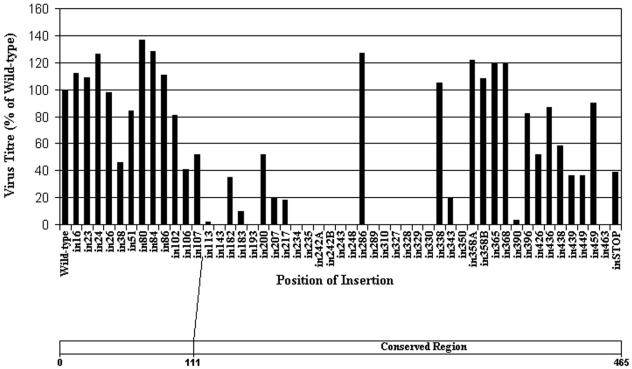
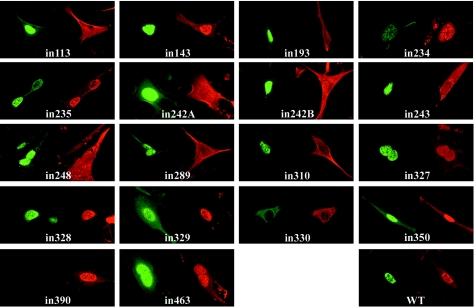
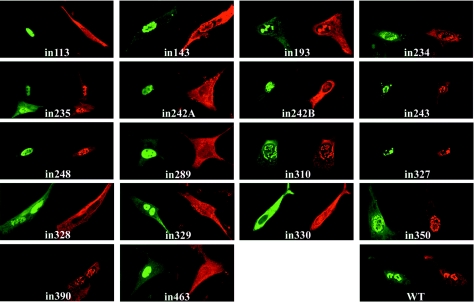
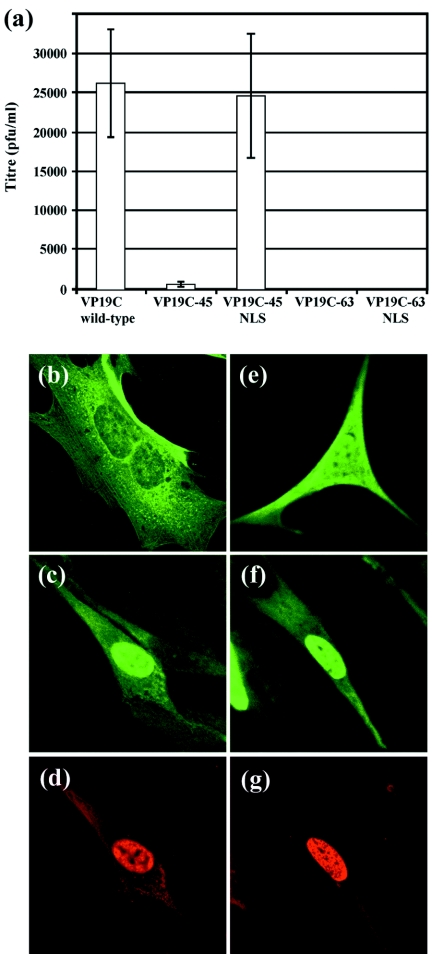
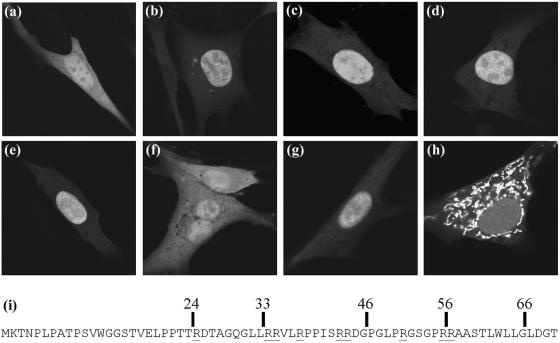
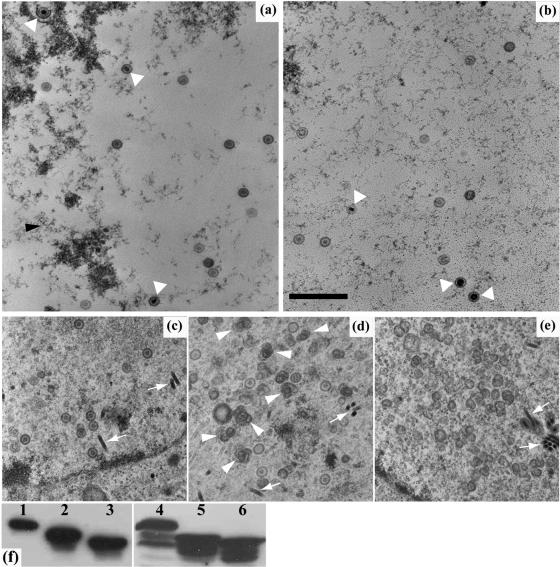
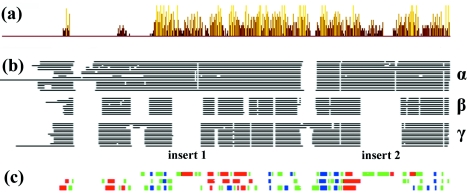
Herpes simplex virus type 1 (HSV-1) capsids have an icosahedral structure with capsomers formed by the major capsid protein, VP5, linked in groups of three by distinctive structures called triplexes. Triplexes are heterotrimers formed by two proteins in a 1:2 stoichiometry. The single-copy protein is called VP19C, and the dimeric protein is VP23. We have carried out insertional and deletional mutagenesis on VP19C and have examined the effects of the mutations on virus growth and capsid assembly. Insertional mutagenesis showed that the N-terminal approximately 100 amino acids of the protein, which correspond to a region that is poorly conserved among herpesviruses, are insensitive to disruption and that insertions into the rest of the protein had various effects on virus growth. Some, but not all, severely disabled mutants were compromised in the ability to bind VP23 or VP5. Analysis of deletion mutants revealed the presence of a nuclear localization signal (NLS) near the N terminus of VP19C, and this was mapped to a 33-amino-acid region by fusion of specific sequences to a green fluorescent protein marker. By replacing the endogenous NLS with that from the simian virus 40 large T antigen, we were able to show that the first 45 amino acids of VP19C were not essential for assembly of functional capsids and infectious virus particles. However, removing the first 63 amino acids resulted in formation of aberrant capsids and prevented virus growth, suggesting that the poorly conserved N-terminal sequences have some as-yet-unidentified function.
Figures
References
-
- Bishop, D. H. L. 1992. Baculovirus expression vectors. Semin. Virol. 3:253-264.
-
- Chen, D. H., H. Jiang, M. Lee, F. Y. Liu, and Z. H. Zhou. 1999. Three-dimensional visualization of tegument/capsid interactions in the intact human cytomegalovirus. Virology 260:10-16. - PubMed
-
- Clamp, M., J. Cuff, S. M. Searle, and G. J. Barton. 2004. The Jalview Java alignment editor. Bioinformatics 20:426-427. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
