

Host-selected amino acid changes at the sialic acid binding pocket of the parvovirus capsid modulate cell binding affinity and determine virulence
- PMID: 16415031
- PMCID: PMC1346950
- DOI: 10.1128/JVI.80.3.1563-1573.2006
Host-selected amino acid changes at the sialic acid binding pocket of the parvovirus capsid modulate cell binding affinity and determine virulence
Abstract
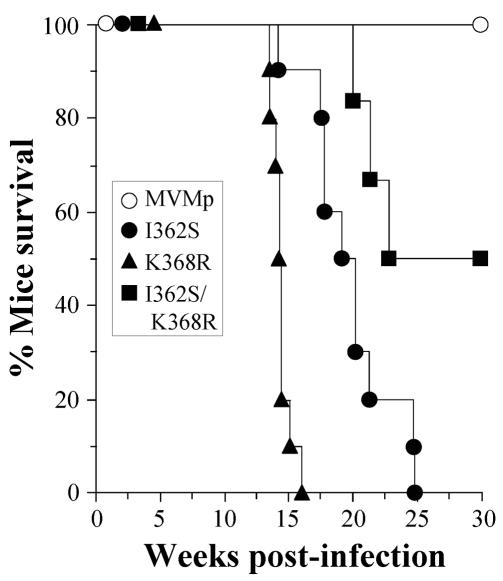
The role of receptor recognition in the emergence of virulent viruses was investigated in the infection of severe combined immunodeficient (SCID) mice by the apathogenic prototype strain of the parvovirus minute virus of mice (MVMp). Genetic analysis of isolated MVMp viral clones (n = 48) emerging in mice, including lethal variants, showed only one of three single changes (V325M, I362S, or K368R) in the common sequence of the two capsid proteins. As was found for the parental isolates, the constructed recombinant viruses harboring the I362S or the K368R single substitutions in the capsid sequence, or mutations at both sites, showed a large-plaque phenotype and lower avidity than the wild type for cells in the cytotoxic interaction with two permissive fibroblast cell lines in vitro and caused a lethal disease in SCID mice when inoculated by the natural oronasal route. Significantly, the productive adsorption of MVMp variants carrying any of the three mutations selected through parallel evolution in mice showed higher sensitivity to the treatment of cells by neuraminidase than that of the wild type, indicating a lower affinity of the viral particle for the sialic acid component of the receptor. Consistent with this, the X-ray crystal structure of the MVMp capsids soaked with sialic acid (N-acetyl neuraminic acid) showed the sugar allocated in the depression at the twofold axis of symmetry (termed the dimple), immediately adjacent to residues I362 and K368, which are located on the wall of the dimple, and approximately 22 A away from V325 in a threefold-related monomer. This is the first reported crystal structure identifying an infectious receptor attachment site on a parvovirus capsid. We conclude that the affinity of the interactions of sialic-acid-containing receptors with residues at or surrounding the dimple can evolutionarily regulate parvovirus pathogenicity and adaptation to new hosts.
Figures
Similar articles
-
Evolution to pathogenicity of the parvovirus minute virus of mice in immunodeficient mice involves genetic heterogeneity at the capsid domain that determines tropism.J Virol. 2008 Feb;82(3):1195-203. doi: 10.1128/JVI.01692-07. Epub 2007 Nov 28. J Virol. 2008. PMID: 18045943 Free PMC article.
-
Virulent variants emerging in mice infected with the apathogenic prototype strain of the parvovirus minute virus of mice exhibit a capsid with low avidity for a primary receptor.J Virol. 2005 Sep;79(17):11280-90. doi: 10.1128/JVI.79.17.11280-11290.2005. J Virol. 2005. PMID: 16103180 Free PMC article.
-
Identification of the sialic acid structures recognized by minute virus of mice and the role of binding affinity in virulence adaptation.J Biol Chem. 2006 Sep 1;281(35):25670-7. doi: 10.1074/jbc.M604421200. Epub 2006 Jul 5. J Biol Chem. 2006. PMID: 16822863
-
Structural determinants of tissue tropism and in vivo pathogenicity for the parvovirus minute virus of mice.J Virol. 2005 Sep;79(17):10931-43. doi: 10.1128/JVI.79.17.10931-10943.2005. J Virol. 2005. PMID: 16103145 Free PMC article.
-
Characterization of the Adeno-Associated Virus 1 and 6 Sialic Acid Binding Site.J Virol. 2016 May 12;90(11):5219-5230. doi: 10.1128/JVI.00161-16. Print 2016 Jun 1. J Virol. 2016. PMID: 26962225 Free PMC article.
Cited by
-
Oncolytic H-1 Parvovirus Enters Cancer Cells through Clathrin-Mediated Endocytosis.Viruses. 2020 Oct 21;12(10):1199. doi: 10.3390/v12101199. Viruses. 2020. PMID: 33096814 Free PMC article.
-
Complex and Dynamic Interactions between Parvovirus Capsids, Transferrin Receptors, and Antibodies Control Cell Infection and Host Range.J Virol. 2018 Jun 13;92(13):e00460-18. doi: 10.1128/JVI.00460-18. Print 2018 Jul 1. J Virol. 2018. PMID: 29695427 Free PMC article.
-
Atomic Resolution Structure of the Oncolytic Parvovirus LuIII by Electron Microscopy and 3D Image Reconstruction.Viruses. 2017 Oct 30;9(11):321. doi: 10.3390/v9110321. Viruses. 2017. PMID: 29084163 Free PMC article.
-
Oncolytic H-1 parvovirus binds to sialic acid on laminins for cell attachment and entry.Nat Commun. 2021 Jun 22;12(1):3834. doi: 10.1038/s41467-021-24034-7. Nat Commun. 2021. PMID: 34158478 Free PMC article.
-
AMDV Vaccine: Challenges and Perspectives.Viruses. 2021 Sep 14;13(9):1833. doi: 10.3390/v13091833. Viruses. 2021. PMID: 34578415 Free PMC article. Review.
References
-
- Agbandje, M., R. McKenna, M. G. Rossmann, M. L. Strassheim, and C. R. Parrish. 1993. Structure determination of feline panleukopenia virus empty particles. Proteins 16:155-171. - PubMed
-
- Agbandje-McKenna, M., A. L. Llamas-Sainz, F. Wang, P. Tattersall, and M. G. Rossmann. 1998. Functional implications of the structure of the murine parvovirus, minute virus of mice. Structure 6:1369-1381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
