

Tudor, MBT and chromo domains gauge the degree of lysine methylation
- PMID: 16415788
- PMCID: PMC1456902
- DOI: 10.1038/sj.embor.7400625
Tudor, MBT and chromo domains gauge the degree of lysine methylation
Abstract
The post-translational modification of histones regulates many cellular processes, including transcription, replication and DNA repair. A large number of combinations of post-translational modifications are possible. This cipher is referred to as the histone code. Many of the enzymes that lay down this code have been identified. However, so far, few code-reading proteins have been identified. Here, we describe a protein-array approach for identifying methyl-specific interacting proteins. We found that not only chromo domains but also tudor and MBT domains bind to methylated peptides from the amino-terminal tails of histones H3 and H4. Binding specificity observed on the protein-domain microarray was corroborated using peptide pull-downs, surface plasma resonance and far western blotting. Thus, our studies expose tudor and MBT domains as new classes of methyl-lysine-binding protein modules, and also demonstrates that protein-domain microarrays are powerful tools for the identification of new domain types that recognize histone modifications.
Figures
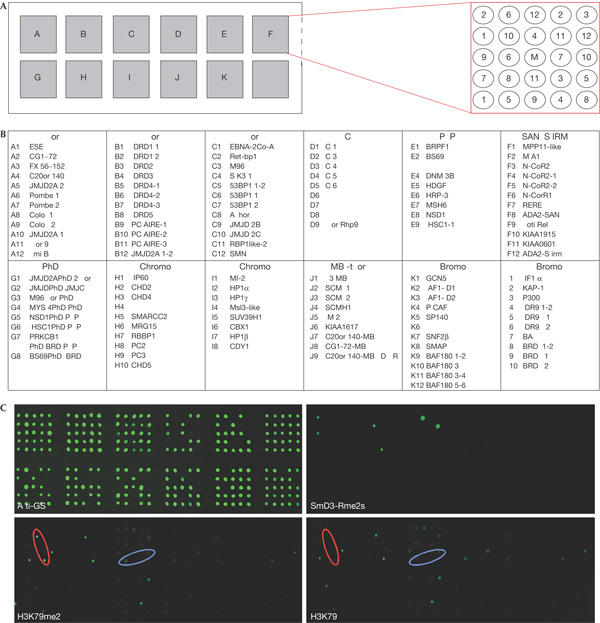

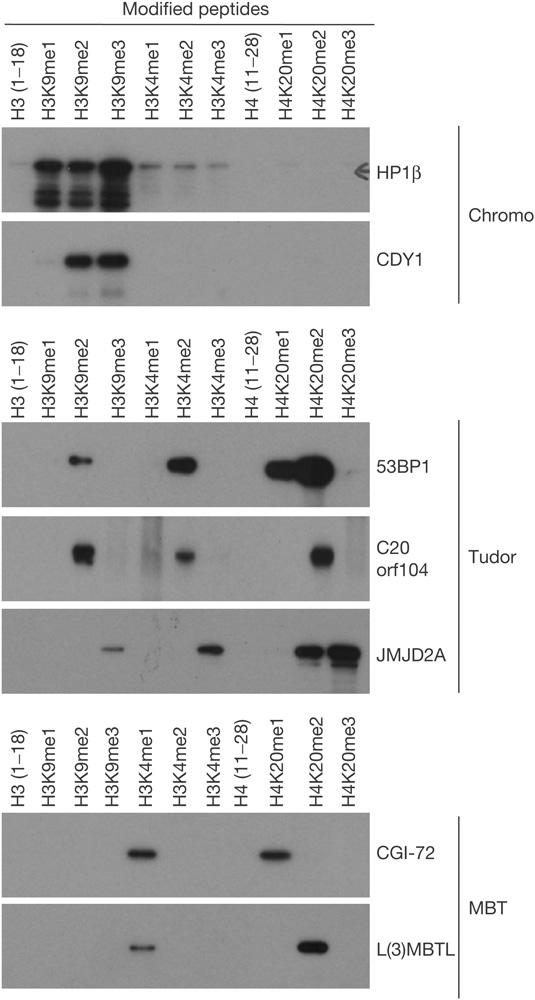
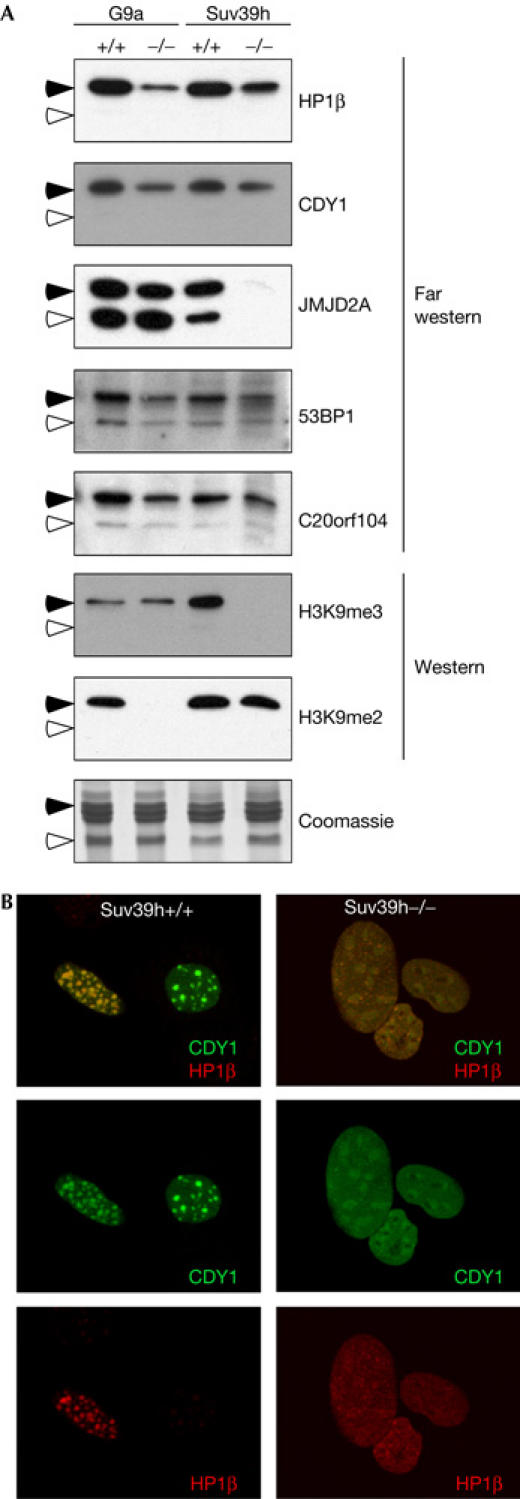
References
-
- Aletta JM, Cimato TR, Ettinger MJ (1998) Protein methylation: a signal event in post-translational modification. Trends Biochem Sci 23: 89–91 - PubMed
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T (2001) Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410: 120–124 - PubMed
-
- Behrends U et al. (2003) Novel tumor antigens identified by autologous antibody screening of childhood medulloblastoma cDNA libraries. Int J Cancer 106: 244–251 - PubMed
-
- Boccuni P, MacGrogan D, Scandura JM, Nimer SD (2003) The human L(3)MBT polycomb group protein is a transcriptional repressor and interacts physically and functionally with TEL (ETV6). J Biol Chem 278: 15412–15420 - PubMed
-
- Butler AP, Byus CV, Slaga TJ (1986) Phosphorylation of histones is stimulated by phorbol esters in quiescent Reuber H35 hepatoma cells. J Biol Chem 261: 9421–9425 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
