

A role for voltage-gated, but not Ca2+-activated, K+ channels in regulating spontaneous contractile activity in myometrium from virgin and pregnant rats
- PMID: 16415906
- PMCID: PMC1751504
- DOI: 10.1038/sj.bjp.0706644
A role for voltage-gated, but not Ca2+-activated, K+ channels in regulating spontaneous contractile activity in myometrium from virgin and pregnant rats
Abstract
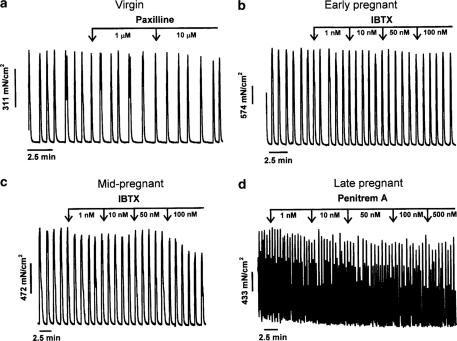
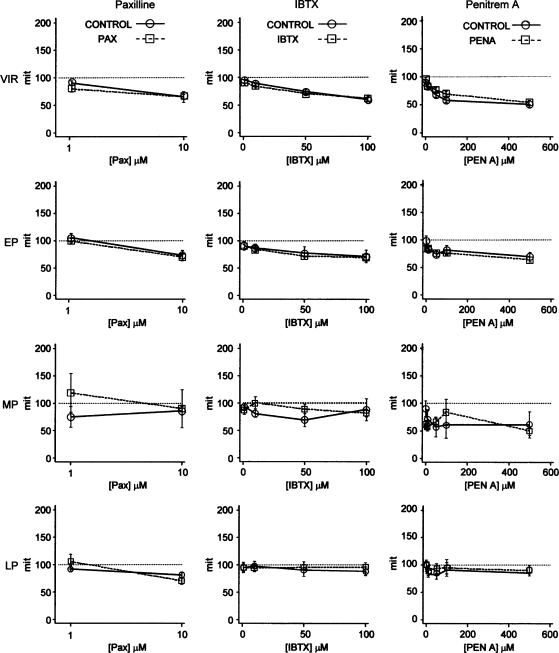
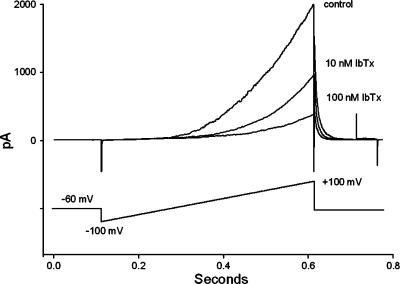
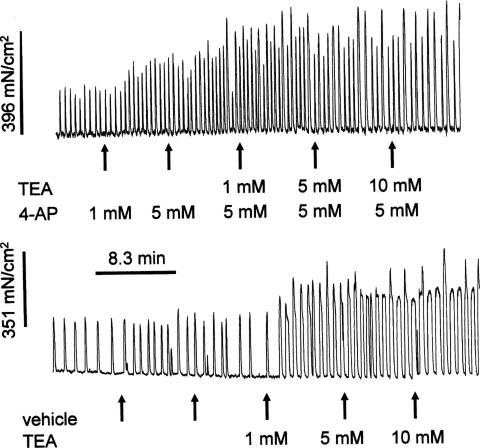
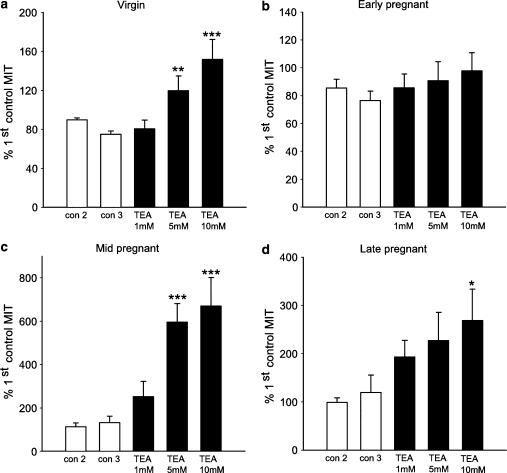
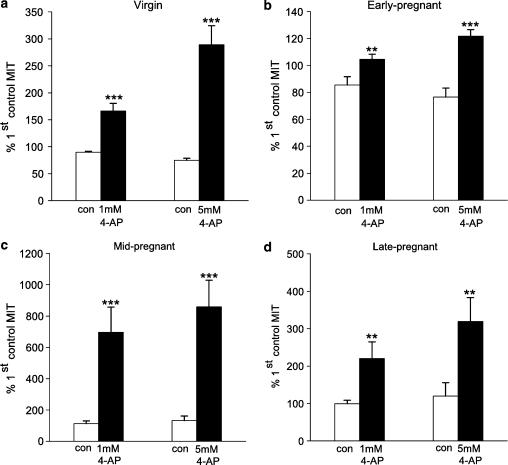
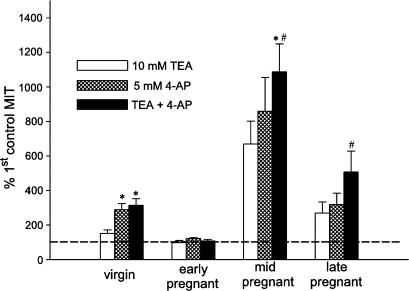
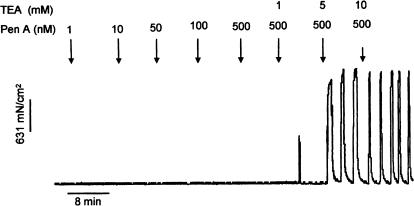
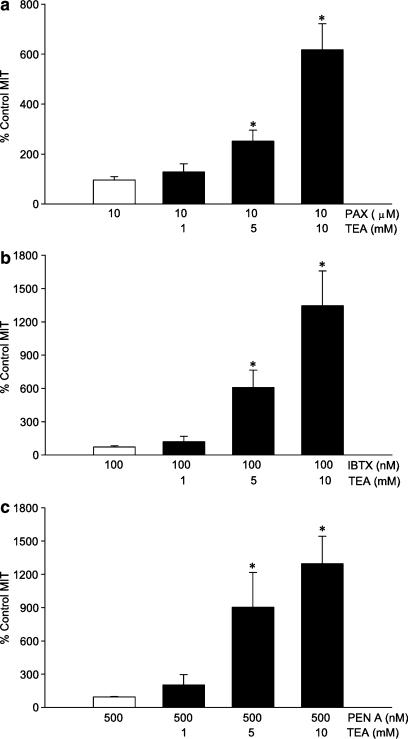
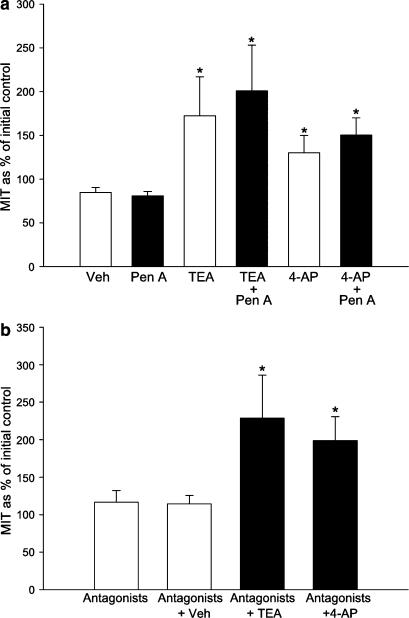
The roles of voltage-gated (K(V)) and large conductance Ca2+-activated K+ (BK(Ca)) channels in regulating basal contractility in myometrial smooth muscle are unresolved. The aim of this study was to determine the effects of inhibition of these channels on spontaneous rhythmic contraction in myometrial strips from four groups of rats: nonpregnant and during early (day 7), mid- (day 14), and late (day 21) pregnancy. BK(Ca) channels were inhibited using iberiotoxin (1-100 nM), paxilline (1-10 microM) or penitrem A (1-500, or 3000 nM); K(V) channels were inhibited using tetraethylammonium (TEA; 1-10 mM) and/or 4-aminopyridine (4-AP; 1-5 mM). Contractility was assessed as mean integral tension (MIT). Time/vehicle controls were also performed. None of the selective BK(Ca) channel inhibitors significantly affected contractility in myometrial strips from either nonpregnant or pregnant animals. 4-AP caused concentration-dependent increases in MIT in myometrium in all four groups. TEA (5 and 10 mM) significantly increased MIT in myometrium from nonpregnant, and mid- and late pregnant rats, but not in myometrium from early pregnant rats. TEA and 4-AP still caused an increase in MIT following treatment with 3000 nM penitrem A or a combination of propranolol, phentolamine, atropine (all 1 microM) and capsaicin (10 microM) in myometrial strips from nonpregnant rats. These results indicate that whereas BK(Ca) channels play little or no part in controlling basal rhythmicity in rat myometrium, K(V) channels appear to play a crucial role in this regard, especially during mid- and late pregnancy.
Figures
References
-
- ADEAGBO A.S.O. 1-Ethyl-2-benzimidazolinone stimulates endothelial KCa channels and nitric oxide formation in rat mesenteric vessels. Eur. J. Pharmacol. 1999;379:151–159. - PubMed
-
- ANWER K., OBERTI C., PEREZ G.J., PEREZ-REYES N., MCDOUGALL J.K., MONGA M., SANBORN B.M., STEFANI E., TORO L. Calcium-activated K+ channels as modulators of human myometrial contractile activity. Am. J. Physiol. 1993;265:C976–C985. - PubMed
-
- ANWER K., TORO L., OBERTI C., STEFANI E., SANBORN B.M. Ca2+-activated K+ channels in pregnant rat myometrium: modulation by a beta-adrenergic agent. Am. J. Physiol. 1992;263:C1049–C1056. - PubMed
-
- BAI X., BUGG G.J., GREENWOOD S.L., GLAZIER J.D., SIBLEY C.P., BAKER P.N., TAGGART M.J., FYFE G.K. Expression of TASK and TREK, two-pore domain K+ channels, in human myometrium. Reproduction. 2005;129:525–530. - PubMed
-
- BRADLEY K.K., BUXTON I.L., BARBER J.E., MCGAW T., BRADLEY M.E. Nitric oxide relaxes human myometrium by a cGMP-independent mechanism. Am. J. Physiol. 1998;275:C1668–C1673. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
