

Molecular characterization of geminivirus-derived small RNAs in different plant species
- PMID: 16421273
- PMCID: PMC1342034
- DOI: 10.1093/nar/gkj447
Molecular characterization of geminivirus-derived small RNAs in different plant species
Abstract
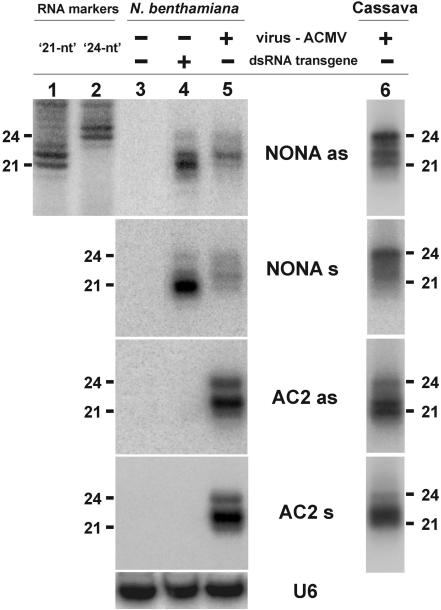
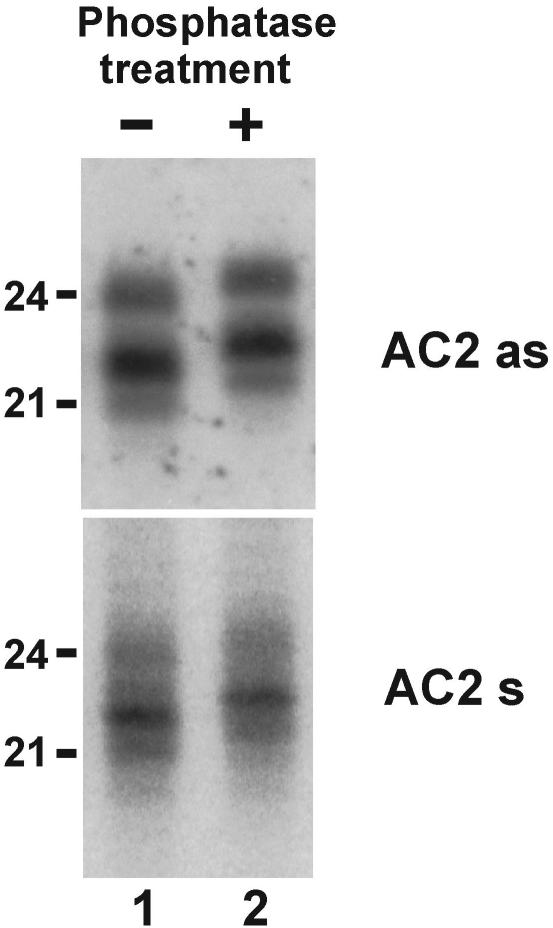
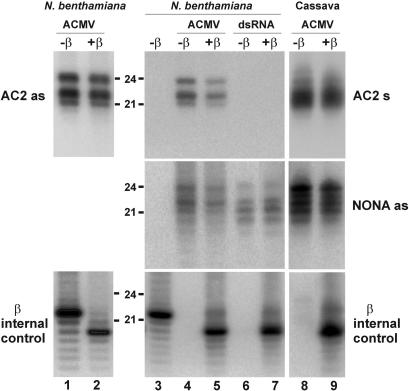
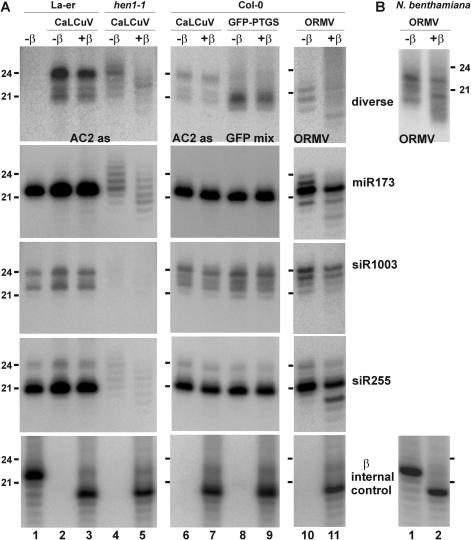
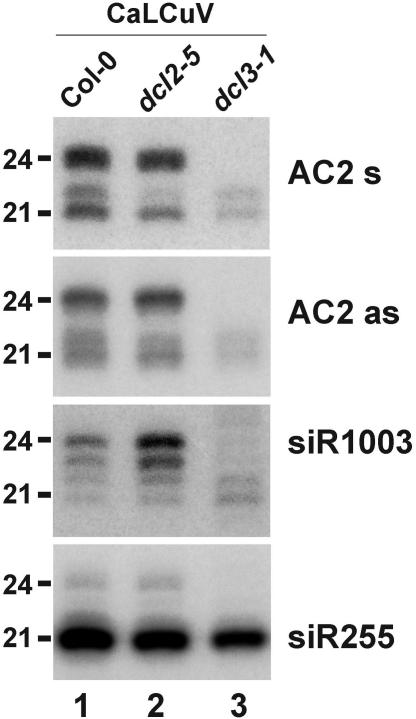
DNA geminiviruses are thought to be targets of RNA silencing. Here, we characterize small interfering (si) RNAs-the hallmarks of silencing-associated with Cabbage leaf curl begomovirus in Arabidopsis and African cassava mosaic begomovirus in Nicotiana benthamiana and cassava. We detected 21, 22 and 24 nt siRNAs of both polarities, derived from both the coding and the intergenic regions of these geminiviruses. Genetic evidence showed that all the 24 nt and a substantial fraction of the 22 nt viral siRNAs are generated by the dicer-like proteins DCL3 and DCL2, respectively. The viral siRNAs were 5' end phosphorylated, as shown by phosphatase treatments, and methylated at the 3'-nucleotide, as shown by HEN1 miRNA methylase-dependent resistance to beta-elimination. Similar modifications were found in all types of endogenous and transgene-derived siRNAs tested, but not in a major fraction of siRNAs from a cytoplasmic RNA tobamovirus. We conclude that several distinct silencing pathways are involved in DNA virus-plant interactions.
Figures
References
-
- Bartel D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Meins F., Jr, Si-Ammour A., Blevins T. RNA silencing systems and their relevance to plant development. Annu. Rev. Cell Dev. Biol. 2005;21:297–318. - PubMed
-
- Baulcombe D. RNA silencing. Trends Biochem. Sci. 2005;30:290–293. - PubMed
-
- Zilberman D., Cao X., Johansen L.K., Xie Z., Carrington J.C., Jacobsen S.E. Role of Arabidopsis ARGONAUTE4 in RNA-directed DNA methylation triggered by inverted repeats. Curr. Biol. 2004;14:1214–1220. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
