

The role of striatum in initiation and execution of learned action sequences in rats
- PMID: 16421321
- PMCID: PMC6675371
- DOI: 10.1523/JNEUROSCI.3883-05.2006
The role of striatum in initiation and execution of learned action sequences in rats
Abstract
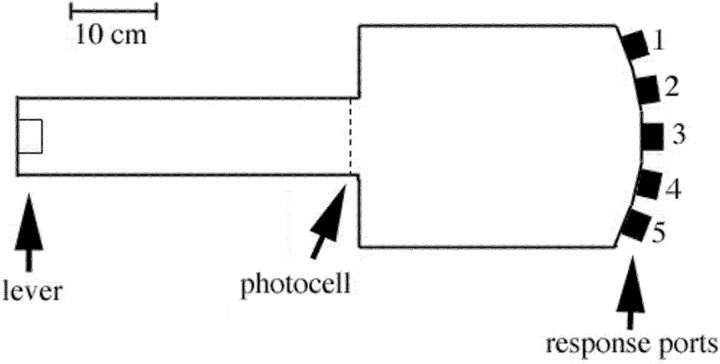
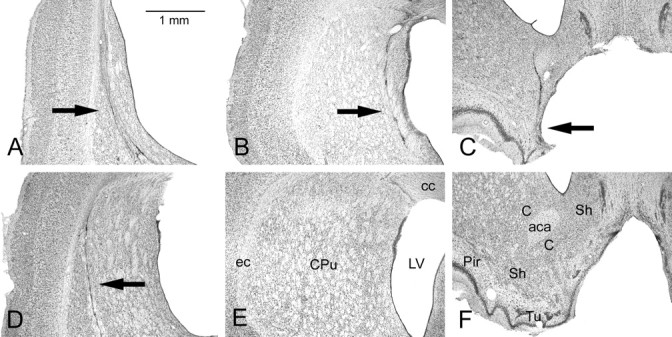
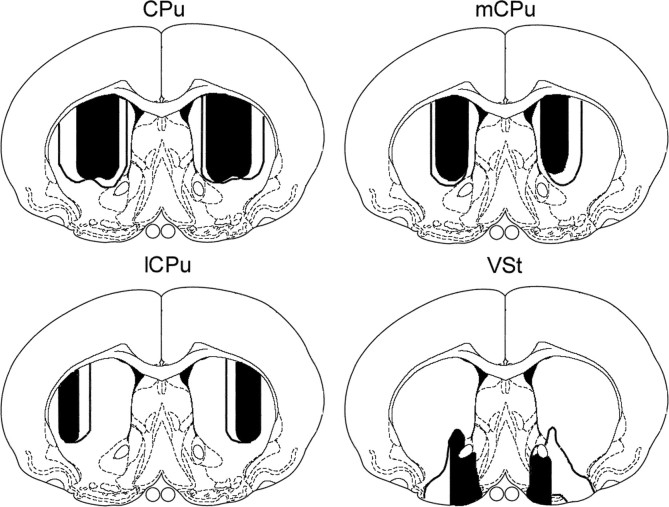
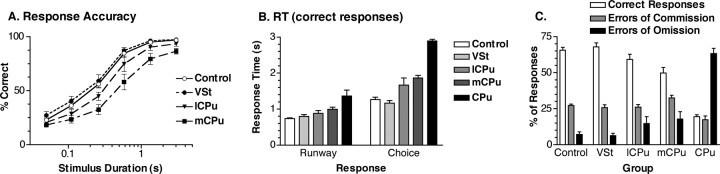
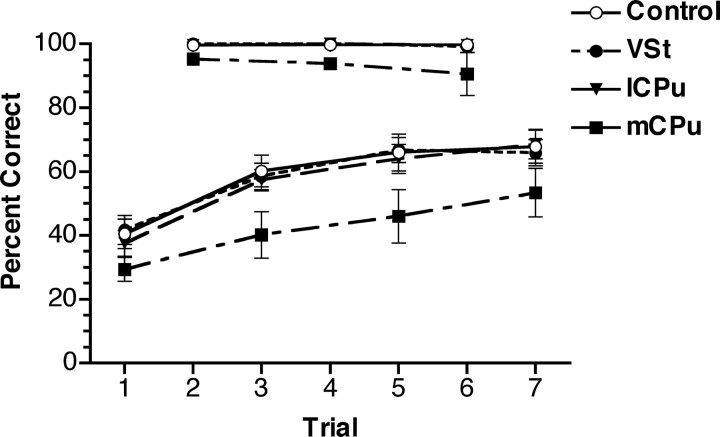
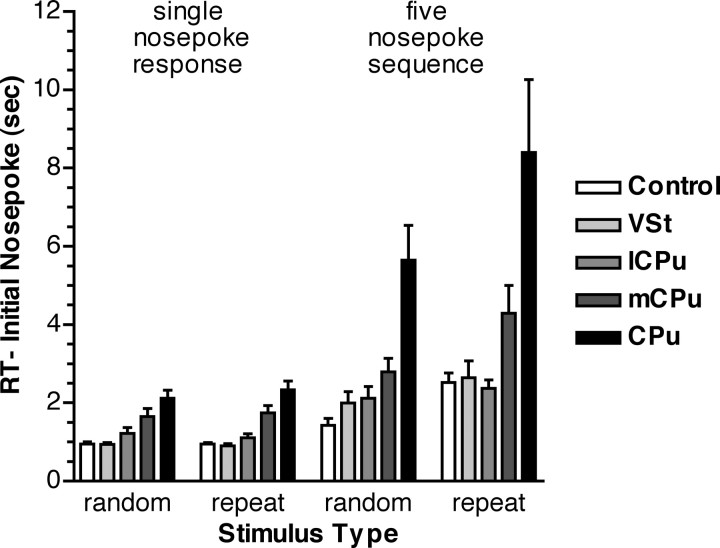
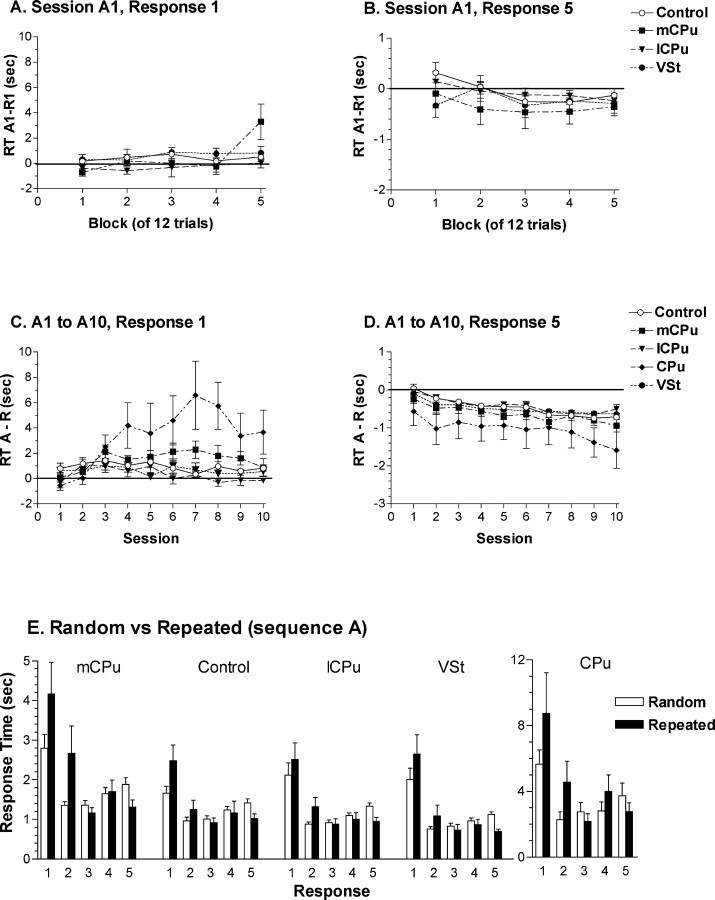
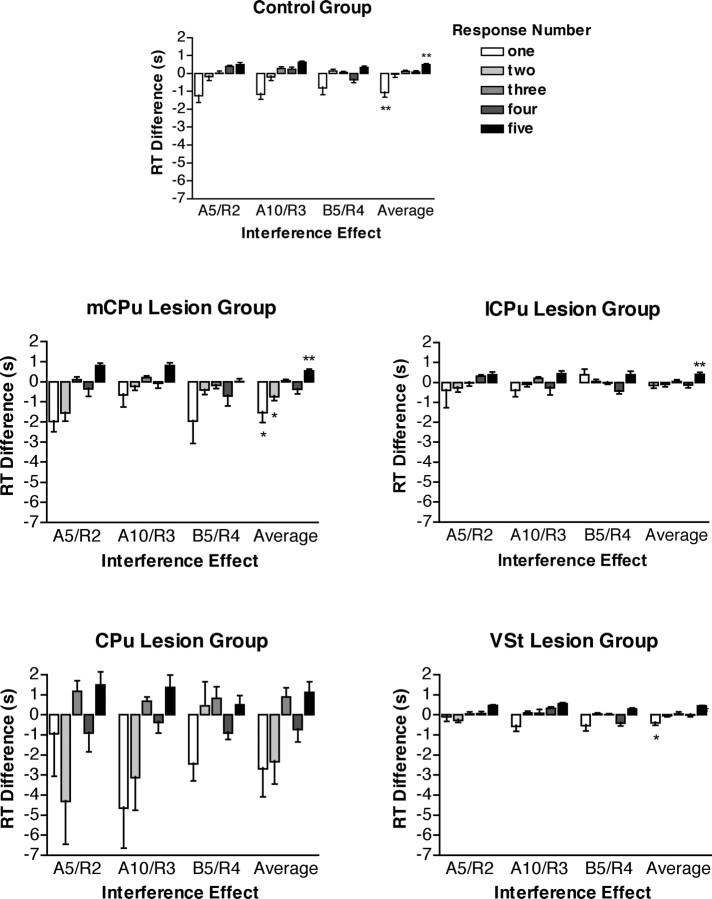
To understand the role of striatum in motor sequence learning, we trained rats to perform a series of tasks measuring speed and accuracy of responding to luminance cues presented as discriminative stimuli for single nose pokes or for sequences of nose pokes in a serial reaction time task. Habit (stimulus-response) learning was measured by comparing performances when stimuli were repeated (predictable) with when they were selected randomly (unpredictable). Sequences had defined start and end points and were limited to five nose pokes to minimize chunking. When sequences were repeated, response time (RT) increased for nose pokes initiating the sequence and decreased for nose pokes completing it. These effects developed incrementally across sessions, consistent with the time course of habit learning. Medial (mCPu), lateral, and complete (CPu) caudate-putamen lesions affected speed and accuracy of single nose poke responses, confirming the role of these areas in guiding responses with external sensory stimuli. None of these lesions affected the short-term increase in accuracy observed when single nose poke responses were repeated. Both mCPu and CPu lesions increased RTs for initiating sequential responses, effects that were exacerbated across sessions in which specific sequences were repeated. None of the lesions affected the gradual decrease in RT for nose pokes completing repeated sequences. Correlational analyses confirmed the relationship between the extent of dorsal striatal damage and the ability to respond to brief luminance cues and to initiate learned sequences. These results provide evidence implicating dorsal striatum in higher-level organizational aspects of learning reflected in planning that precedes the execution of learned action sequences.
Figures
References
-
- Bailey KR, Mair RG (2004) Dissociable effects of frontal cortical lesions on measures of visuospatial attention and spatial working memory in the rat. Cereb Cortex 14: 974–985. - PubMed
-
- Bailey KR, Zhang YP, Toupin MM, Mair RG (2004) Premotor and prefrontal cortical lesions affect time to initiate, but not learning-related changes in motor sequence responding. Soc Neurosci Abstr 30: 771.8.
-
- Brown RG, Redondo-Verge L, Chacon JR, Lucas ML, Channon S (2001) Dissociation between intentional and incidental sequence learning in Huntington's disease. Brain 124: 2188–2202. - PubMed
-
- Burk JA, Mair RG (2001) Effects of dorsal and ventral striatal lesions on delayed matching trained with retractable levers. Behav Brain Res 122: 67–78. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources