

TLR agonists abrogate costimulation blockade-induced prolongation of skin allografts
- PMID: 16424185
- PMCID: PMC3916149
- DOI: 10.4049/jimmunol.176.3.1561
TLR agonists abrogate costimulation blockade-induced prolongation of skin allografts
Abstract
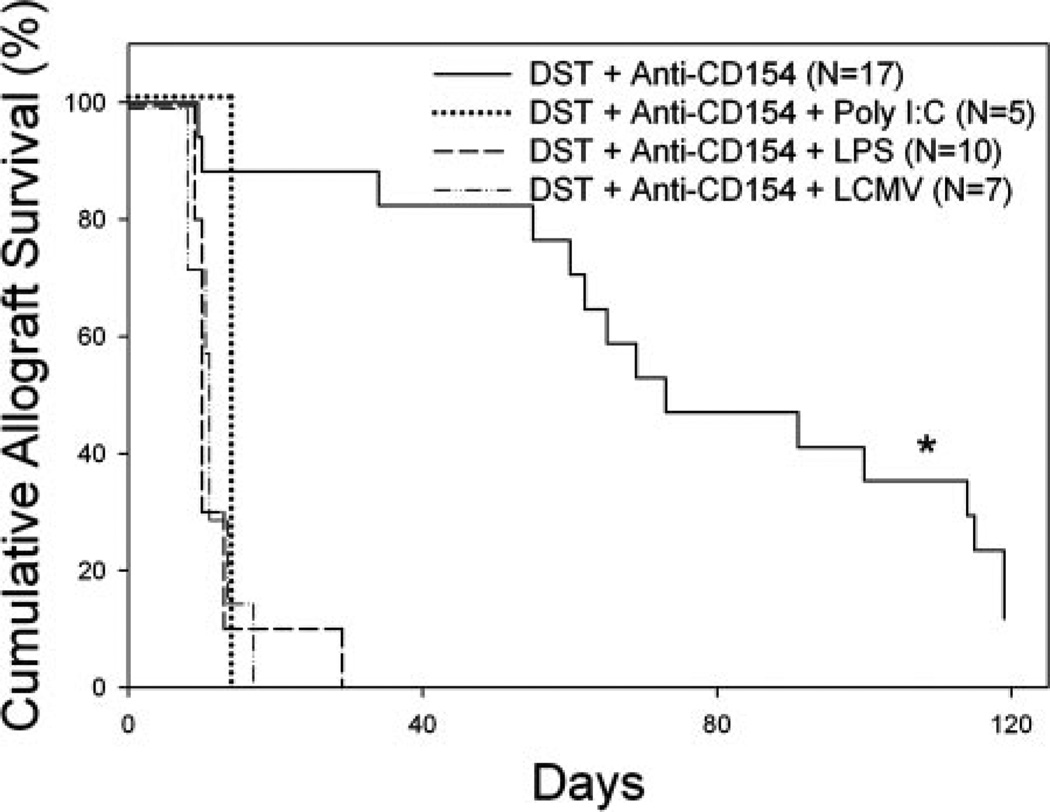
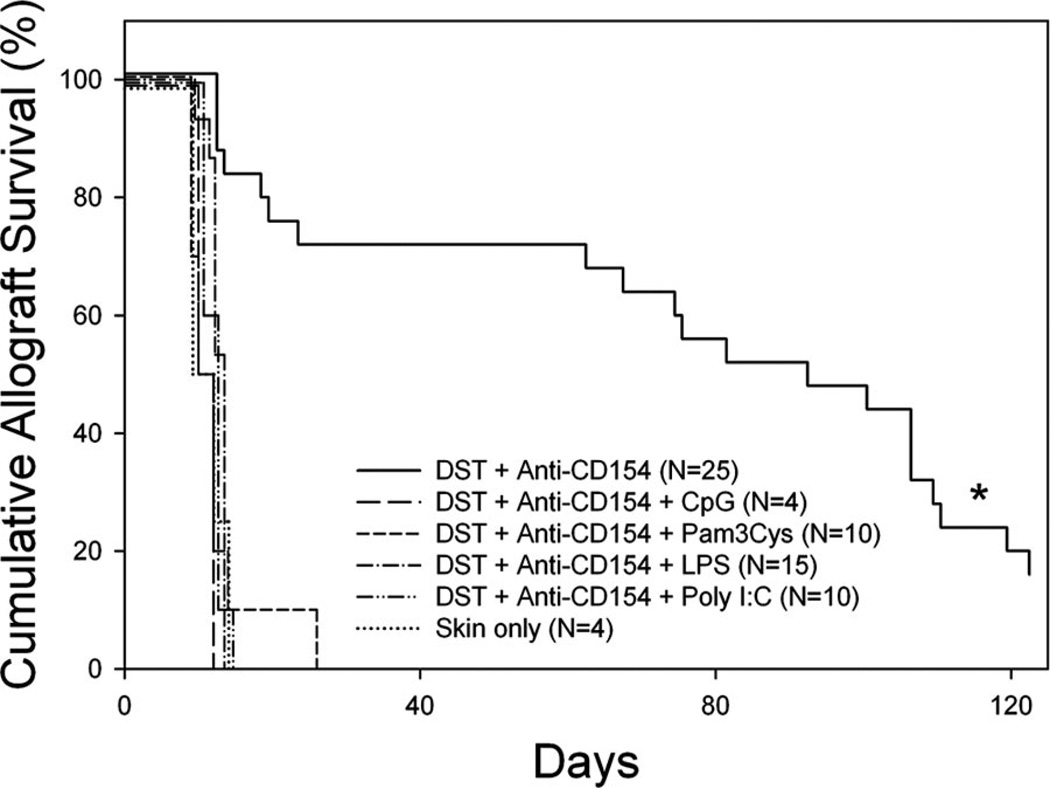
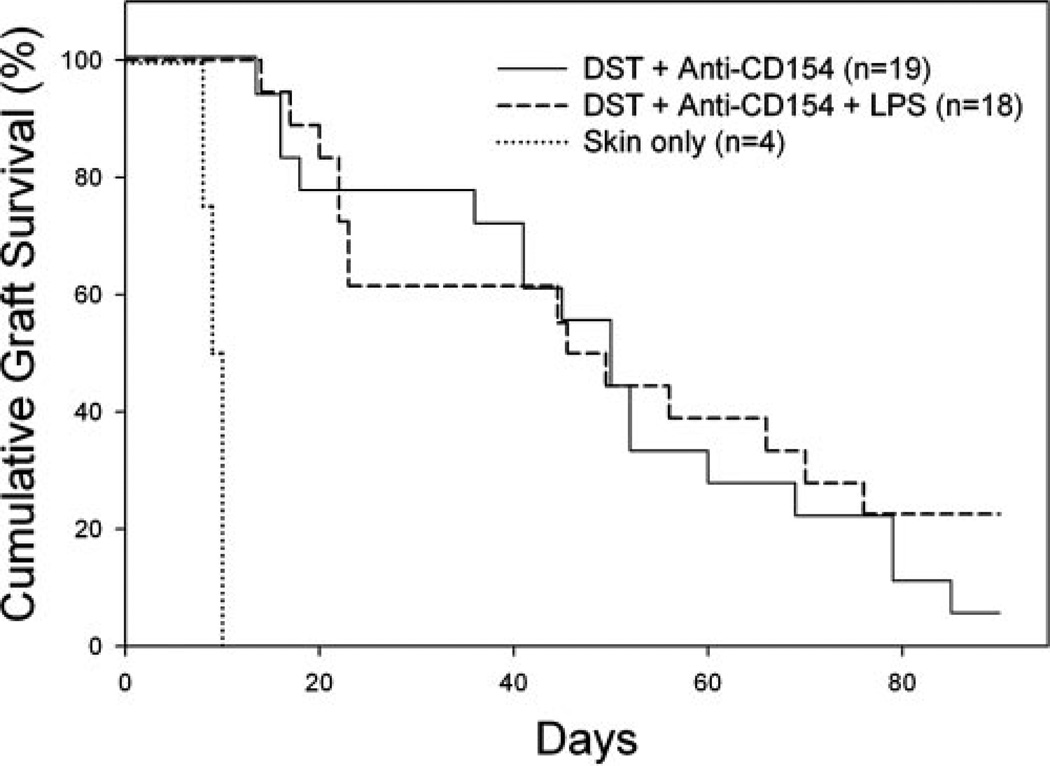
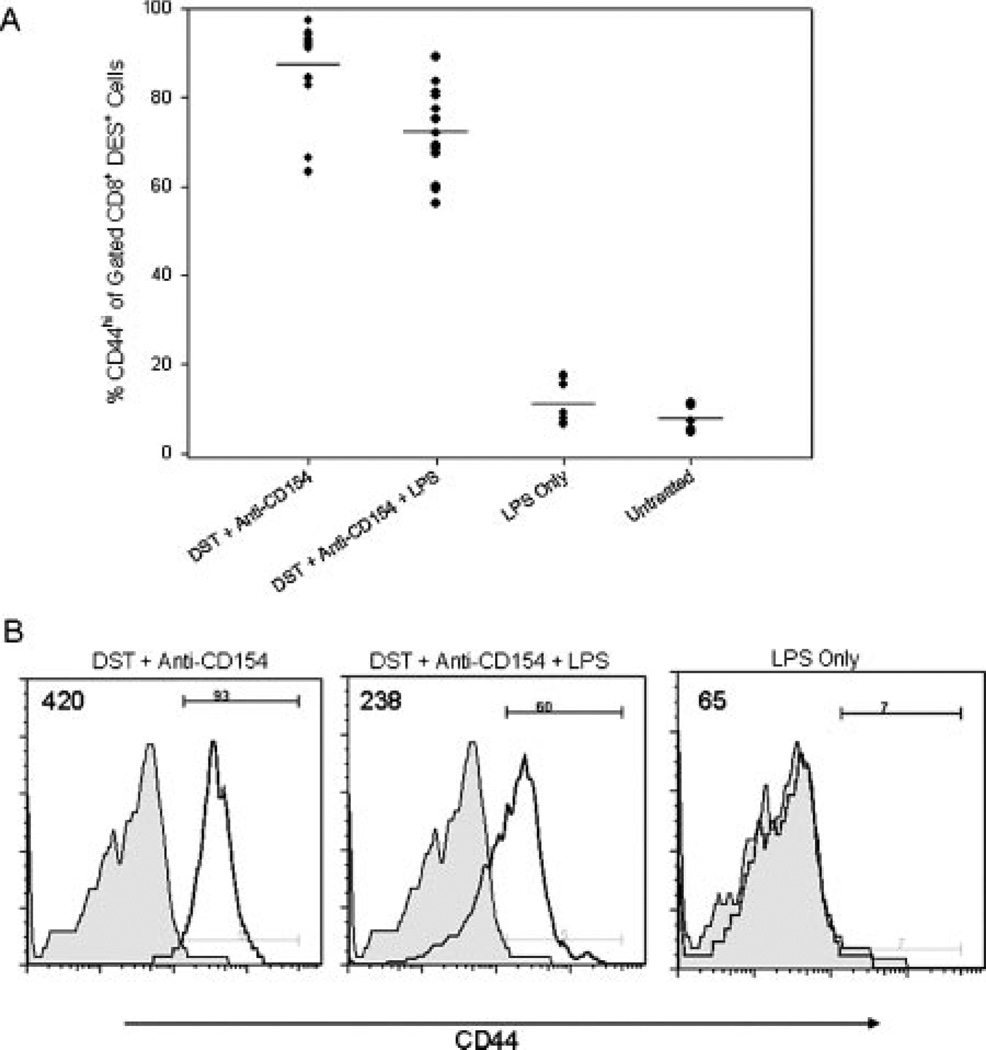
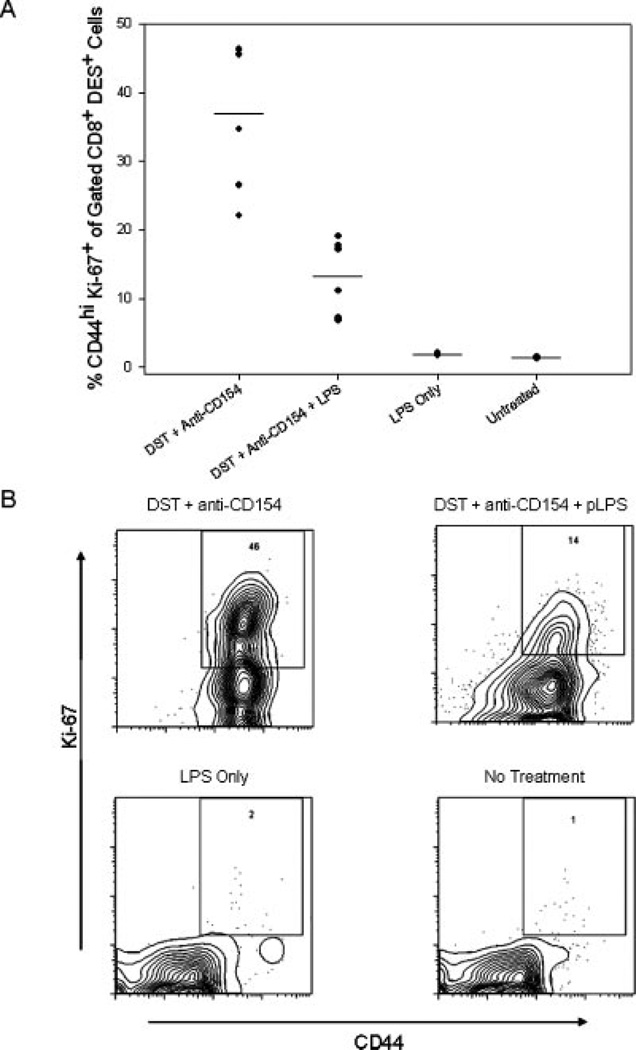
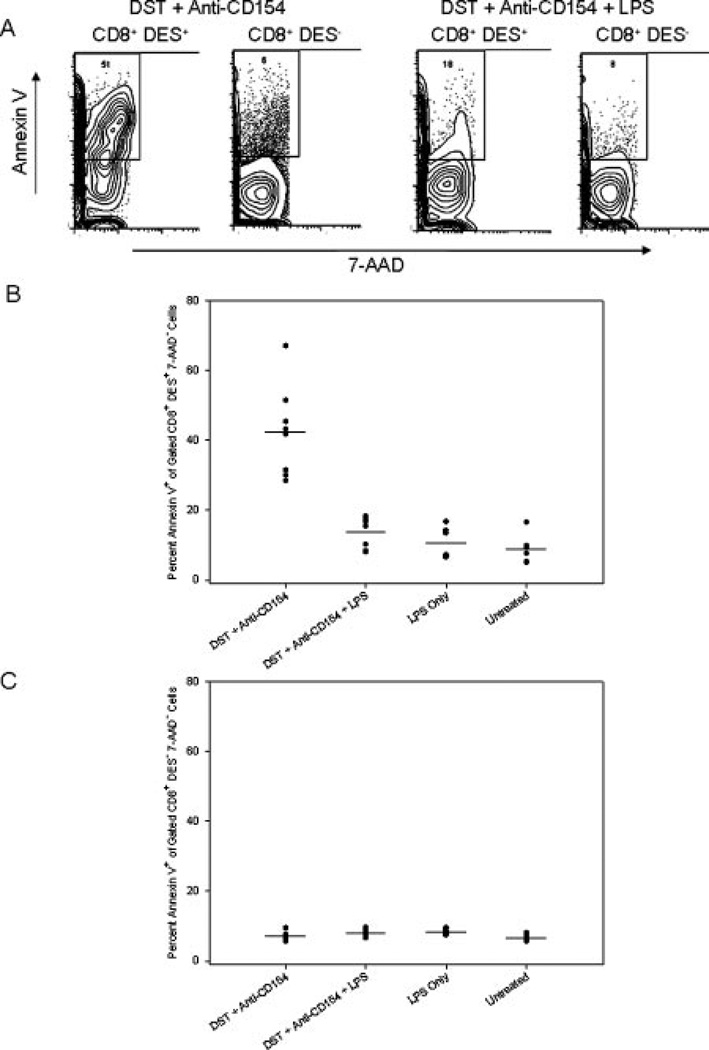
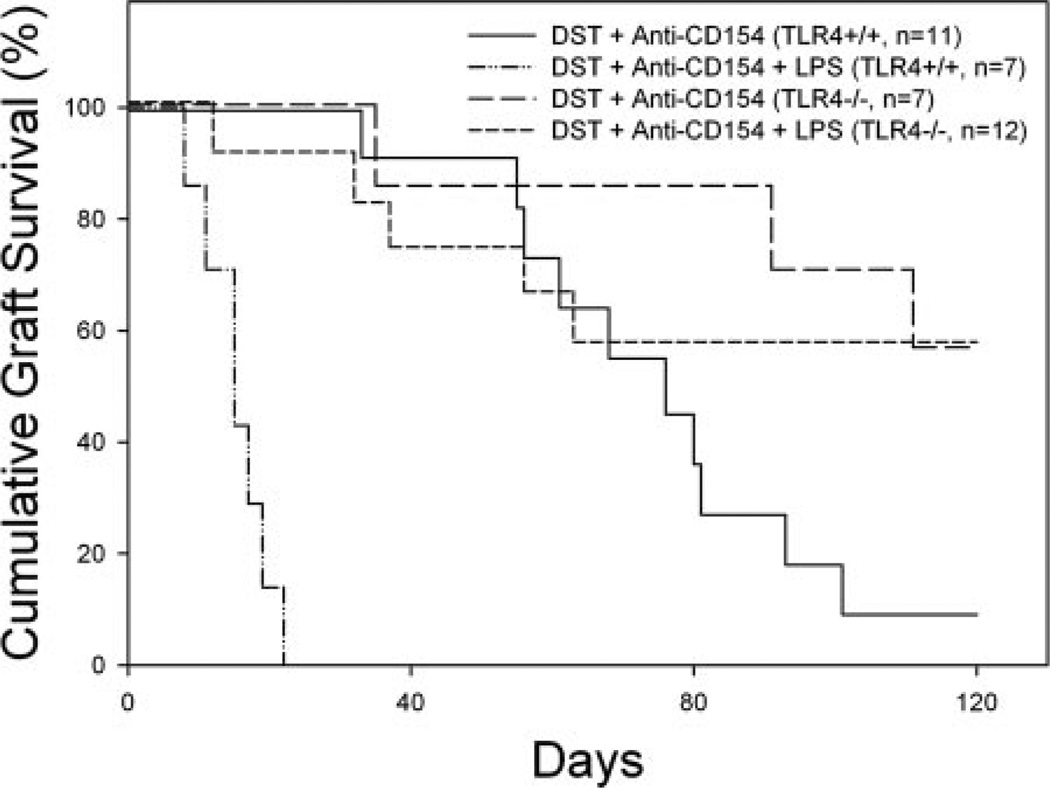
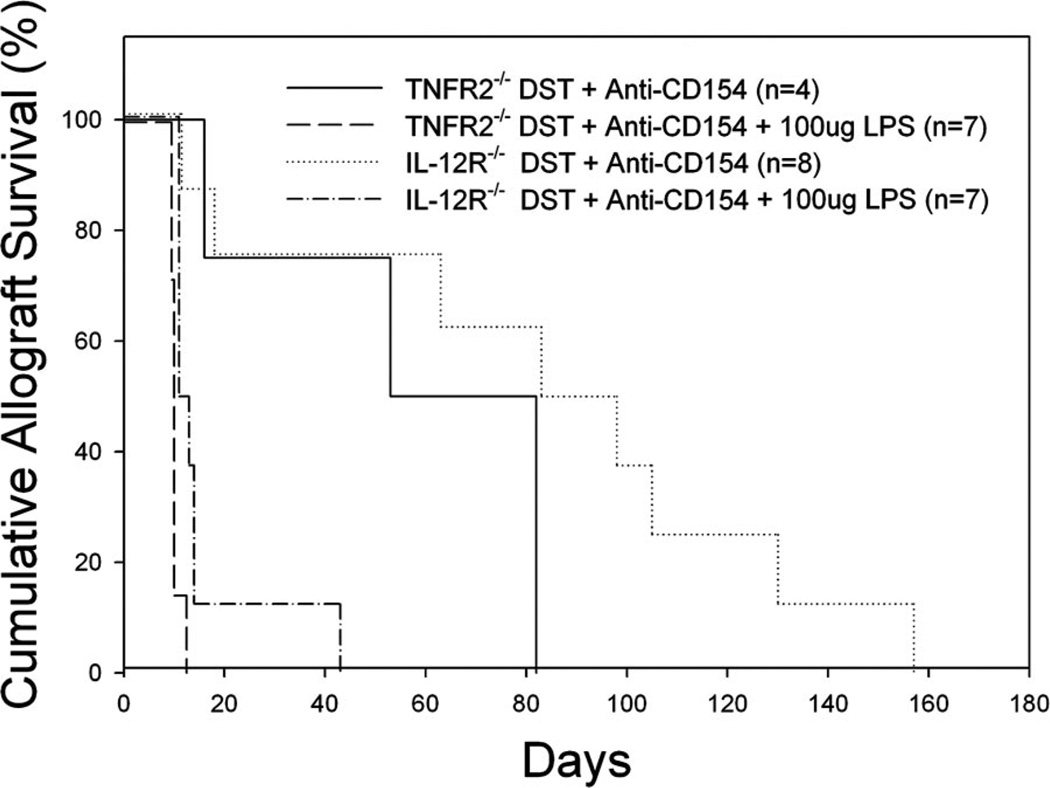
Costimulation blockade protocols are effective in prolonging allograft survival in animal models and are entering clinical trials, but how environmental perturbants affect graft survival remains largely unstudied. We used a costimulation blockade protocol consisting of a donor-specific transfusion and anti-CD154 mAb to address this question. We observed that lymphocytic choriomeningitis virus infection at the time of donor-specific transfusion and anti-CD154 mAb shortens allograft survival. Lymphocytic choriomeningitis virus 1) activates innate immunity, 2) induces allo-cross-reactive T cells, and 3) generates virus-specific responses, all of which may adversely affect allograft survival. To investigate the role of innate immunity, mice given costimulation blockade and skin allografts were coinjected with TLR2 (Pam3Cys), TLR3 (polyinosinic:polycytidylic acid), TLR4 (LPS), or TLR9 (CpG) agonists. Costimulation blockade prolonged skin allograft survival that was shortened after coinjection by TLR agonists. To investigate underlying mechanisms, we used "synchimeric" mice which circulate trace populations of anti-H2b transgenic alloreactive CD8+ T cells. In synchimeric mice treated with costimulation blockade, coadministration of all four TLR agonists prevented deletion of alloreactive CD8+ T cells and shortened skin allograft survival. These alloreactive CD8+ T cells 1) expressed the proliferation marker Ki-67, 2) up-regulated CD44, and 3) failed to undergo apoptosis. B6.TNFR2-/- and B6.IL-12R-/- mice treated with costimulation blockade plus LPS also exhibited short skin allograft survival whereas similarly treated B6.CD8alpha-/- and TLR4-/- mice exhibited prolonged allograft survival. We conclude that TLR signaling abrogates the effects of costimulation blockade by preventing alloreactive CD8+ T cell apoptosis through a mechanism not dependent on TNFR2 or IL-12R signaling.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures
References
-
- Rickels MR, Schutta MH, Markmann JF, Barker CF, Naji A, Teff KL. β-cell function following human islet transplantation for type 1 diabetes. Diabetes. 2005;54:100–106. - PubMed
-
- Street CN, Lakey JRT, Shapiro AMJ, Imes S, Rajotte RV, Ryan EA, Lyon JG, Kin T, Avila J, Tsujimura T, Korbutt GS. Islet graft assessment in the Edmonton protocol-implications for predicting long-term clinical outcome. Diabetes. 2004;53:3107–3114. - PubMed
-
- Shapiro AMJ, Lakey JRT, Ryan EA, Korbutt GS, Toth E, Warnock GL, Kneteman NM, Rajotte RV. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl. J. Med. 2000;343:230–238. - PubMed
-
- Ryan EA, Lakey JRT, Paty BW, Imes S, Korbutt GS, Kneteman NM, Bigam D, Rajotte RV, Shapiro AMJ. Successful islet transplantation-continued insulin reserve provides long-term glycemic control. Diabetes. 2002;51:2148–2157. - PubMed
-
- Teraoka S, Kawai T, Yamaguchi Y, Fujita S, Tojinbara T, Nakajima T, Hayashi T, Nakagawa Y, Fujikawa H, Honda H, et al. Mechanism of the preventive effect of aminobenzoic acid salt on pancreatic β cell toxicity by cyclosporin. Transplant. Proc. 1990;22:863–866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
