

Chromosomes can congress to the metaphase plate before biorientation
- PMID: 16424343
- PMCID: PMC4768465
- DOI: 10.1126/science.1122142
Chromosomes can congress to the metaphase plate before biorientation
Abstract
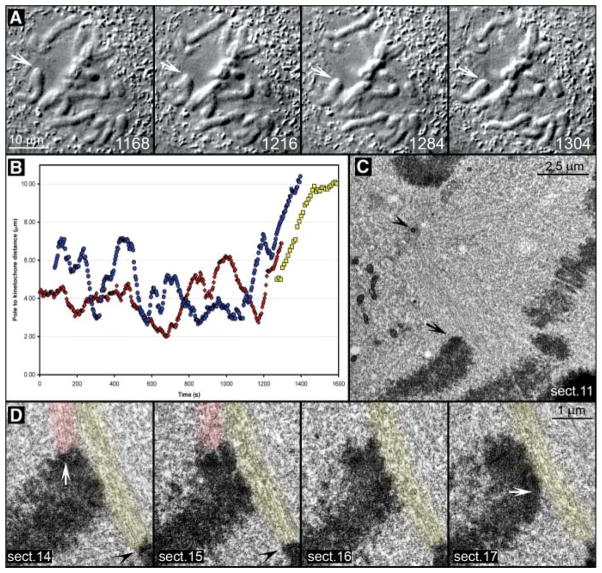
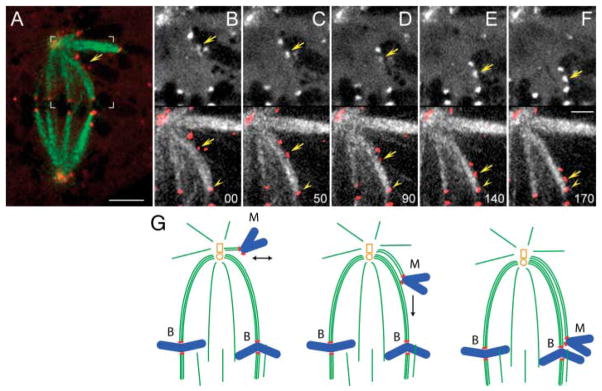
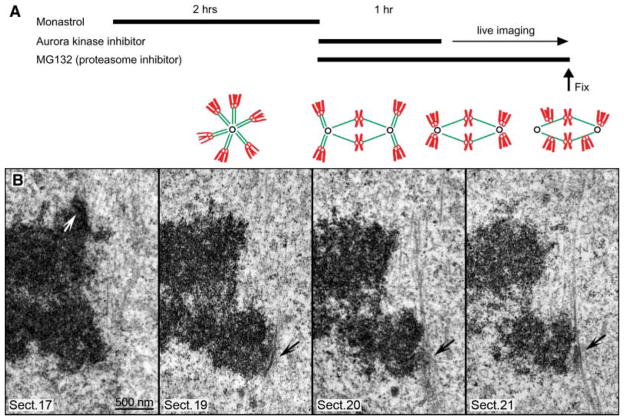
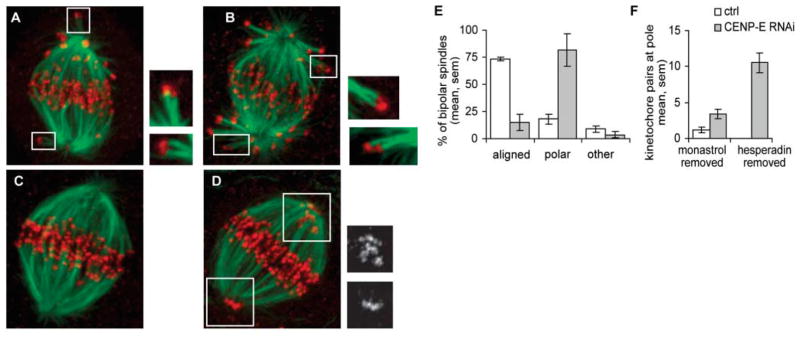
The stable propagation of genetic material during cell division depends on the congression of chromosomes to the spindle equator before the cell initiates anaphase. It is generally assumed that congression requires that chromosomes are connected to the opposite poles of the bipolar spindle ("bioriented"). In mammalian cells, we found that chromosomes can congress before becoming bioriented. By combining the use of reversible chemical inhibitors, live-cell light microscopy, and correlative electron microscopy, we found that monooriented chromosomes could glide toward the spindle equator alongside kinetochore fibers attached to other already bioriented chromosomes. This congression mechanism depended on the kinetochore-associated, plus end-directed microtubule motor CENP-E (kinesin-7).
Figures
Comment in
-
Cell biology. Serving up a plate of chromosomes.Science. 2006 Jan 20;311(5759):343-4. doi: 10.1126/science.1123525. Science. 2006. PMID: 16424328 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
