

Telomere length homeostasis requires that telomerase levels are limiting
- PMID: 16424902
- PMCID: PMC1383536
- DOI: 10.1038/sj.emboj.7600952
Telomere length homeostasis requires that telomerase levels are limiting
Abstract
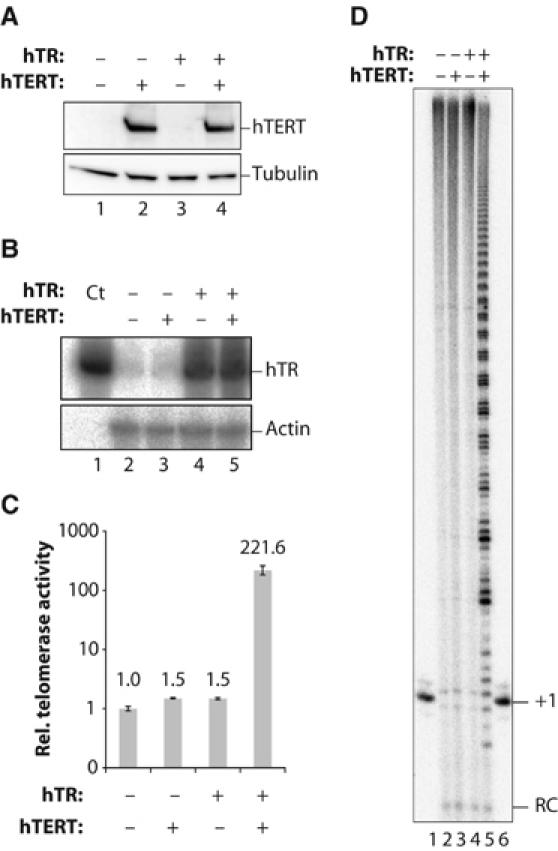
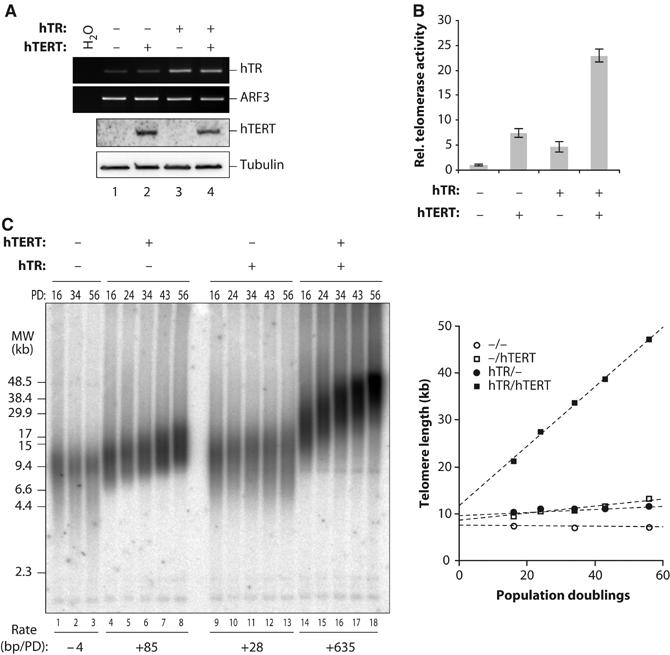
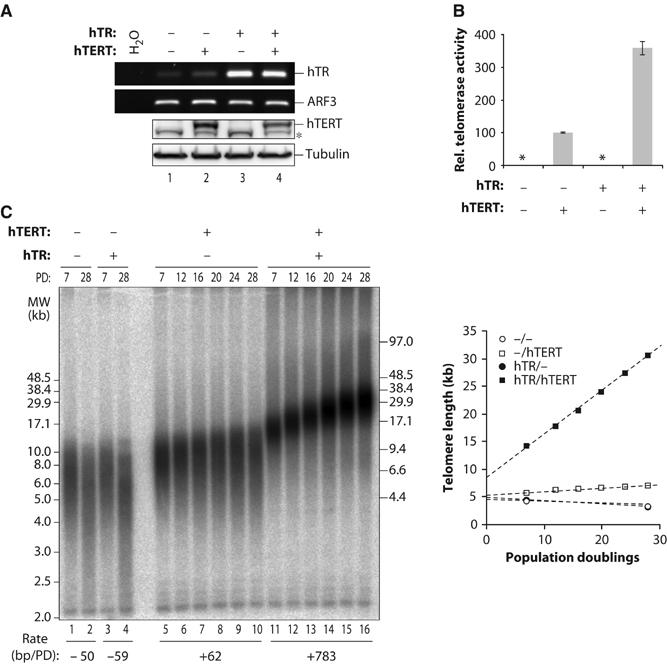
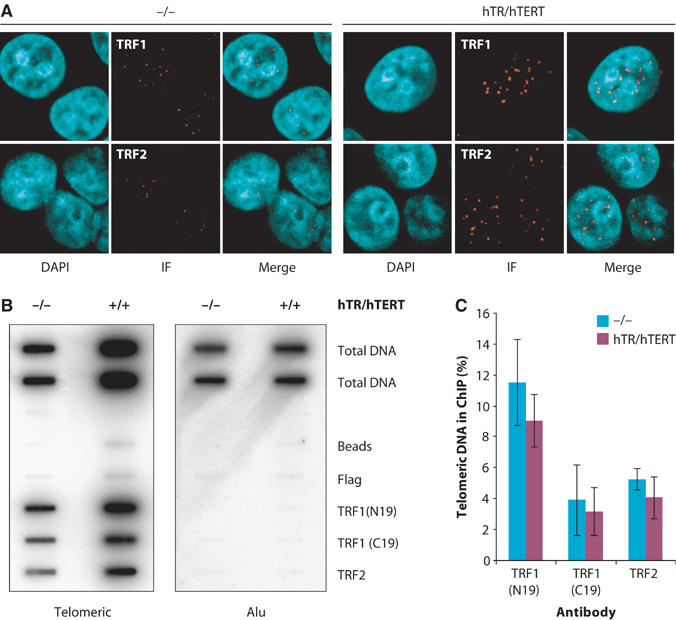
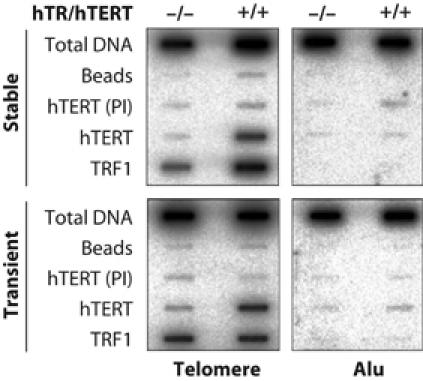
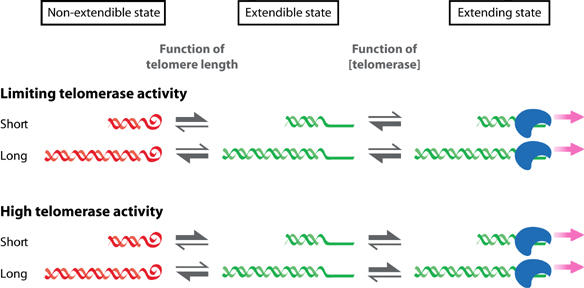
Stabilization of telomere length in germline and highly proliferative human cells is required for long-term survival and for the immortal phenotype of cancer-derived cells. This is achieved through expression of telomerase reverse transcriptase (TERT), which synthesizes telomeric repeats through reverse transcription of its tightly associated RNA template (TR). The telomeric repeat binding factor TRF1 inhibits telomerase at telomeres in cis in a length-dependent manner to achieve telomere length homeostasis. Here we manipulate telomerase activity over a wide range in cancer and primary cells. Concomitant overexpression of TERT and TR was necessary and sufficient to substantially increase telomerase activity. Upon overexpression, more telomerase associated with telomeres and telomeres elongated at a constant rate (up to 0.8 kb/population doubling (PD)) in a length-independent manner. Thus, in less than 50 PDs, the length of telomeres increased 3-8-fold beyond physiological size, while telomere-bound TRF1 and TRF2 increased proportionally to telomere length. Thus, long telomeres do not permanently adopt a structural state that is non-extendible. A low cellular concentration of telomerase is critical to achieve preferential elongation of short telomeres and telomere length homeostasis.
Figures

References
-
- Ancelin K, Brunori M, Bauwens S, Koering CE, Brun C, Ricoul M, Pommier JP, Sabatier L, Gilson E (2002) Targeting assay to study the cis functions of human telomeric proteins: evidence for inhibition of telomerase by TRF1 and for activation of telomere degradation by TRF2. Mol Cell Biol 22: 3474–3487 - PMC - PubMed
-
- Avilion AA, Piatyszek MA, Gupta J, Shay JW, Bacchetti S, Greider CW (1996) Human telomerase RNA and telomerase activity in immortal cell lines and tumor tissues. Cancer Res 56: 645–650 - PubMed
-
- Baur JA, Zou Y, Shay JW, Wright WE (2001) Telomere position effect in human cells. Science 292: 2075–2077 - PubMed
-
- Beattie TL, Zhou W, Robinson MO, Harrington L (1998) Reconstitution of human telomerase activity in vitro. Curr Biol 8: 177–180 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
