

Evidence that the S.cerevisiae Sgs1 protein facilitates recombinational repair of telomeres during senescence
- PMID: 16428246
- PMCID: PMC1342037
- DOI: 10.1093/nar/gkj452
Evidence that the S.cerevisiae Sgs1 protein facilitates recombinational repair of telomeres during senescence
Abstract
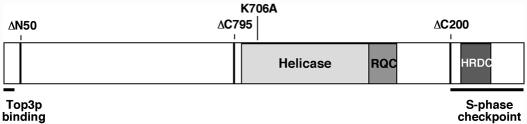
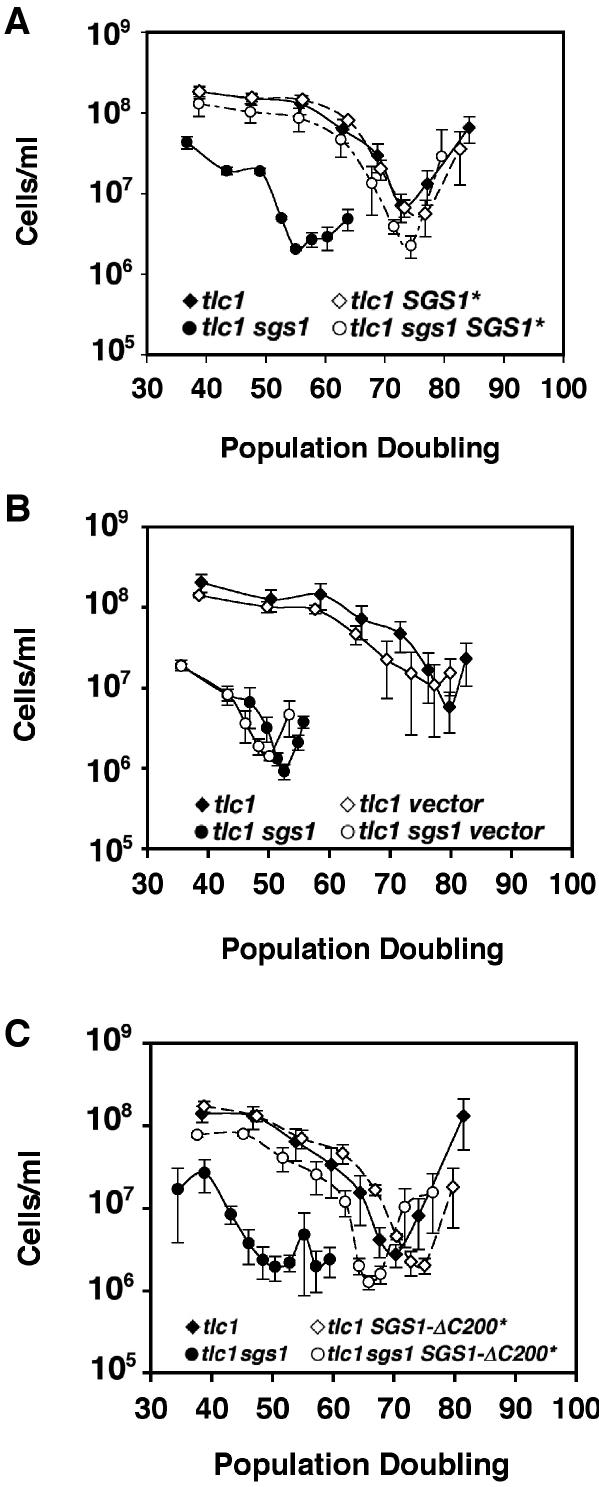
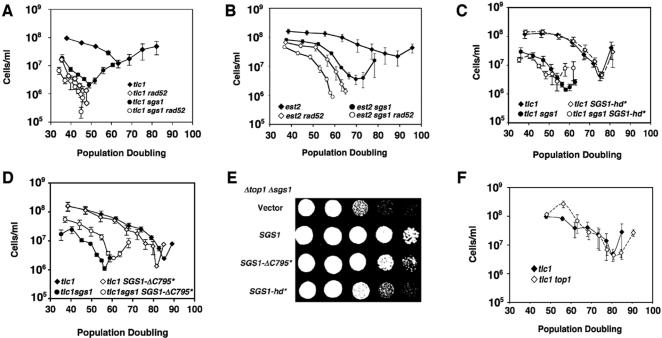
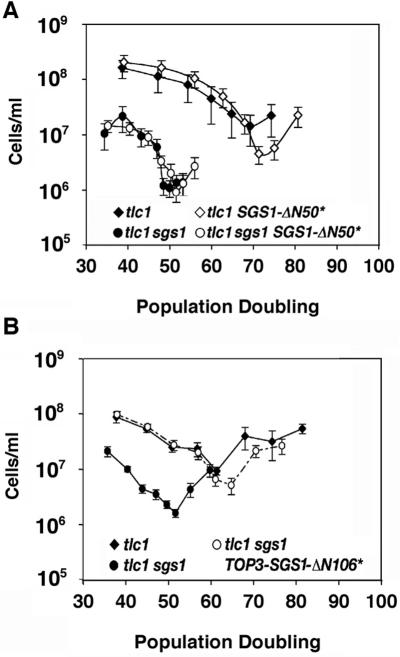
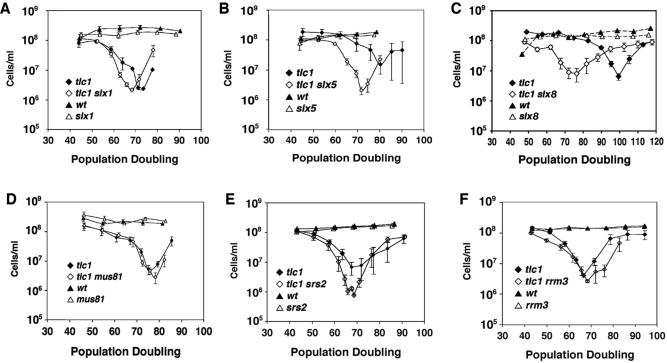
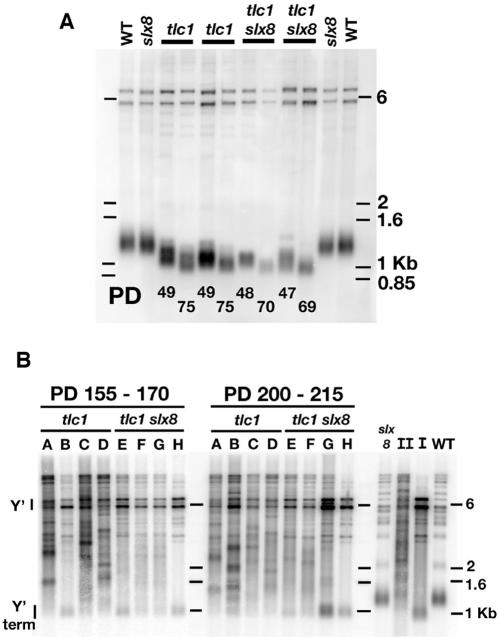
RecQ DNA helicases, including yeast Sgs1p and the human Werner and Bloom syndrome proteins, participate in telomere biology, but the underlying mechanisms are not fully understood. Here, we explore the protein sequences and genetic interactors of Sgs1p that function to slow the senescence of telomerase (tlc1) mutants. We find that the S-phase checkpoint function of Sgs1p is dispensable for preventing rapid senescence, but that Sgs1p sequences required for homologous recombination, including the helicase domain and topoisomerase III interaction domain, are essential. sgs1 and rad52 mutations are epistatic during senescence, indicating that Sgs1p participates in a RAD52-dependent recombinational pathway of telomere maintenance. Several mutations that are synthetically lethal with sgs1 mutation and which individually lead to genome instability, including mus81, srs2, rrm3, slx1 and top1, do not speed the senescence of tlc1 mutants, indicating that the rapid senescence of sgs1 tlc1 mutants is not caused by generic genome instability. However, mutations in SLX5 or SLX8, which encode proteins that function together in a complex that is required for viability in sgs1 mutants, do speed the senescence of tlc1 mutants. These observations further define roles for RecQ helicases and related proteins in telomere maintenance.
Figures
References
-
- Watt P.M., Louis E.J., Borts R.H., Hickson I.D. Sgs1: a eukaryotic homolog of E.coli RecQ that interacts with topoisomerase II in vivo and is required for faithful chromosome segregation. Cell. 1995;81:253–260. - PubMed
-
- Khakhar R.R., Cobb J.A., Bjergbaek L., Hickson I.D., Gasser S.M. RecQ helicases: multiple roles in genome maintenance. Trends Cell Biol. 2003;13:493–501. - PubMed
-
- Hickson I.D. RecQ helicases: caretakers of the genome. Nature Rev. Cancer. 2003;3:169–178. - PubMed
-
- Ellis N.A., Groden J., Ye T.Z., Straughen J., Lennon D.J., Ciocci S., Proytcheva M., German J. The Bloom's syndrome gene product is homologous to RecQ helicases. Cell. 1995;83:655–666. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
