

Identification and characterization of RbmA, a novel protein required for the development of rugose colony morphology and biofilm structure in Vibrio cholerae
- PMID: 16428409
- PMCID: PMC1347326
- DOI: 10.1128/JB.188.3.1049-1059.2006
Identification and characterization of RbmA, a novel protein required for the development of rugose colony morphology and biofilm structure in Vibrio cholerae
Abstract
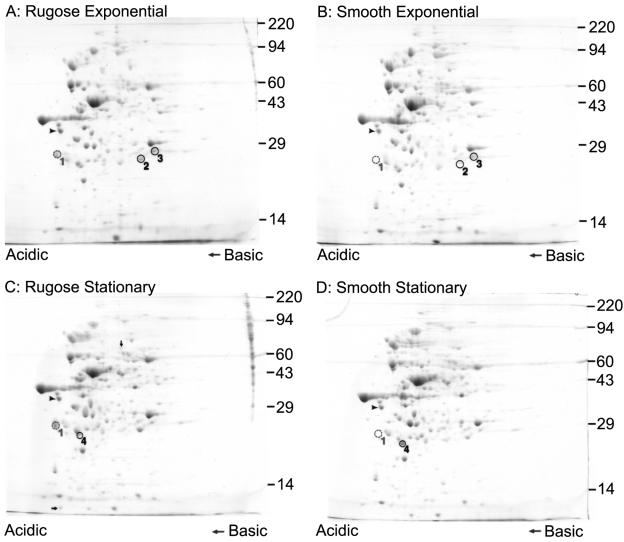
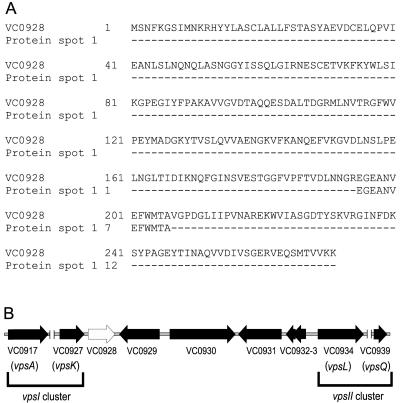
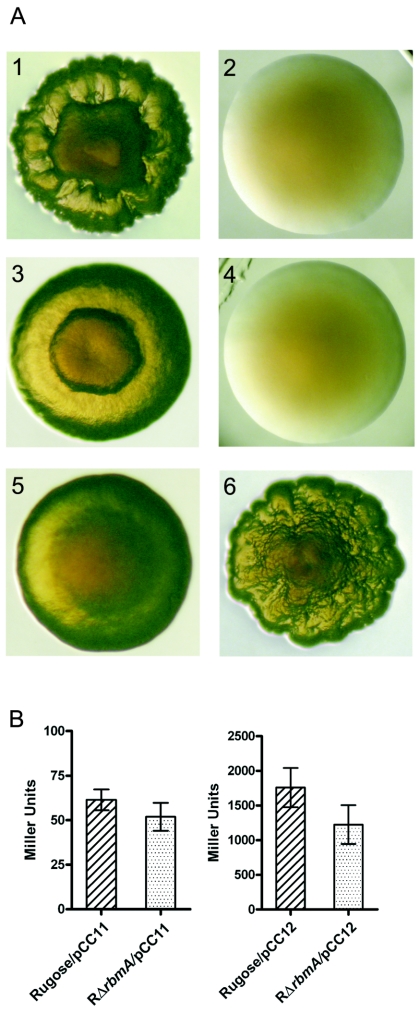
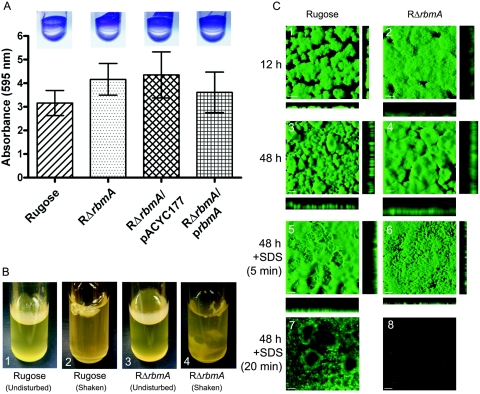
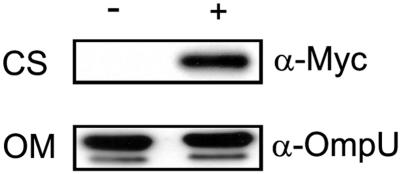
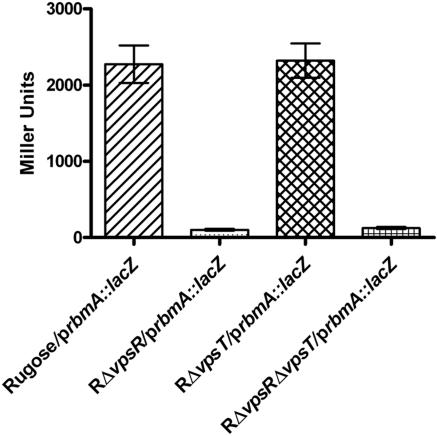
Phase variation between smooth and rugose colony variants of Vibrio cholerae is predicted to be important for the pathogen's survival in its natural aquatic ecosystems. The rugose variant forms corrugated colonies, exhibits increased levels of resistance to osmotic, acid, and oxidative stresses, and has an enhanced capacity to form biofilms. Many of these phenotypes are mediated in part by increased production of an exopolysaccharide termed VPS. In this study, we compared total protein profiles of the smooth and rugose variants using two-dimensional gel electrophoresis and identified one protein that is present at a higher level in the rugose variant. A mutation in the gene encoding this protein, which does not have any known homologs in the protein databases, causes cells to form biofilms that are more fragile and sensitive to sodium dodecyl sulfate than wild-type biofilms. The results indicate that the gene, termed rbmA (rugosity and biofilm structure modulator A), is required for rugose colony formation and biofilm structure integrity in V. cholerae. Transcription of rbmA is positively regulated by the response regulator VpsR but not VpsT.
Figures
References
-
- Ali, A., J. A. Johnson, A. A. Franco, D. J. Metzger, T. D. Connell, J. G. Morris, Jr., and S. Sozhamannan. 2000. Mutations in the extracellular protein secretion pathway genes (eps) interfere with rugose polysaccharide production in and motility of Vibrio cholerae. Infect. Immun. 68:1967-1974. - PMC - PubMed
-
- Bollag, D. M., M. D. Rozycki, and S. J. Edelstein. 1996. Protein methods, 2nd ed., p. 84-85. Eiley-Lis, Inc., New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
