

Inhibition of DNA binding by differential sumoylation of heat shock factors
- PMID: 16428449
- PMCID: PMC1347039
- DOI: 10.1128/MCB.26.3.955-964.2006
Inhibition of DNA binding by differential sumoylation of heat shock factors
Abstract
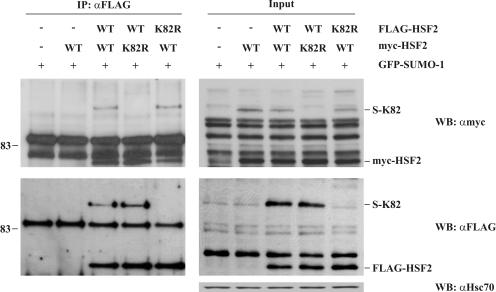
Covalent modification of proteins by the small ubiquitin-related modifier SUMO regulates diverse biological functions. Sumoylation usually requires a consensus tetrapeptide, through which the binding of the SUMO-conjugating enzyme Ubc9 to the target protein is directed. However, additional specificity determinants are in many cases required. To gain insights into SUMO substrate selection, we have utilized the differential sumoylation of highly similar loop structures within the DNA-binding domains of heat shock transcription factor 1 (HSF1) and HSF2. Site-specific mutagenesis in combination with molecular modeling revealed that the sumoylation specificity is determined by several amino acids near the consensus site, which are likely to present the SUMO consensus motif to Ubc9. Importantly, we also demonstrate that sumoylation of the HSF2 loop impedes HSF2 DNA-binding activity, without affecting its oligomerization. Hence, SUMO modification of the HSF2 loop contributes to HSF-specific regulation of DNA binding and broadens the concept of sumoylation in the negative regulation of gene expression.
Figures
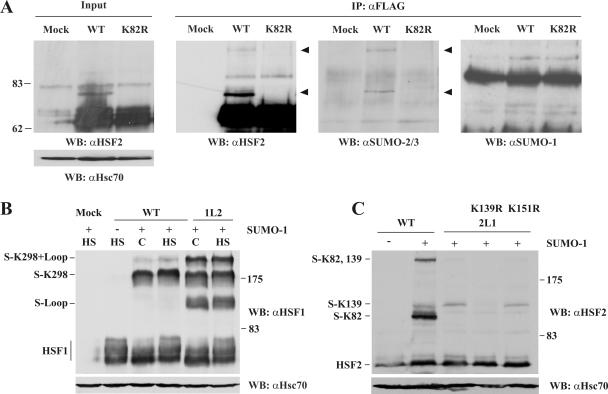
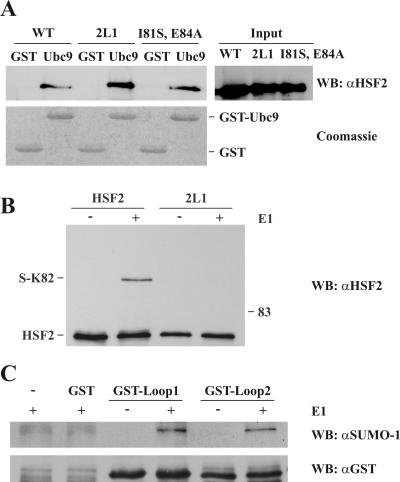
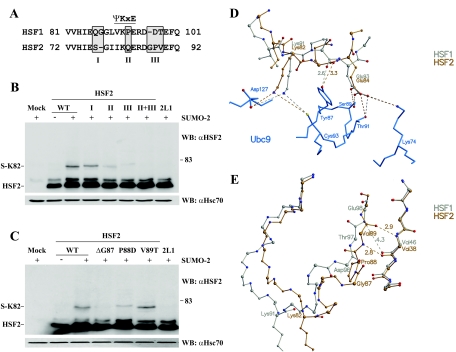
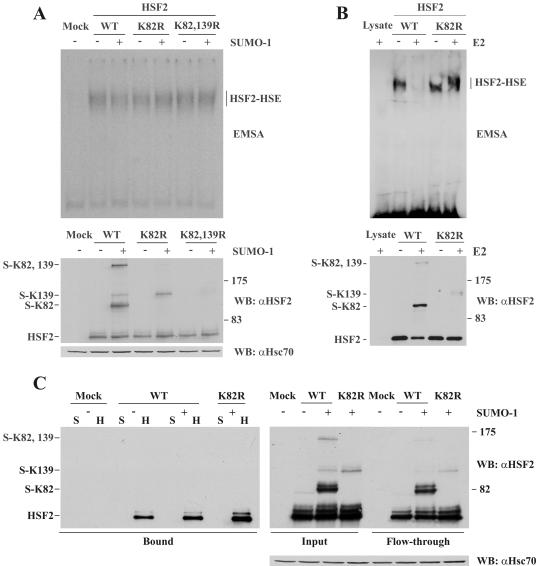
References
-
- Alastalo, T.-P., M. Hellesuo, A. Sandqvist, V. Hietakangas, M. Kallio, and L. Sistonen. 2003. Formation of nuclear stress granules involves HSF2 and coincides with the nucleolar localization of Hsp70. J. Cell Sci. 116:3557-3570. - PubMed
-
- Bernier-Villamor, V., D. A. Sampson, M. J. Matunis, and C. J. Lima. 2002. Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108:345-356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous