

Chloroplastic NAD(P)H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress
- PMID: 16428601
- PMCID: PMC1475475
- DOI: 10.1104/pp.105.070490
Chloroplastic NAD(P)H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress
Abstract
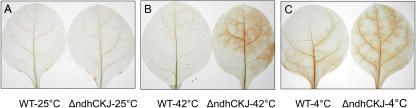
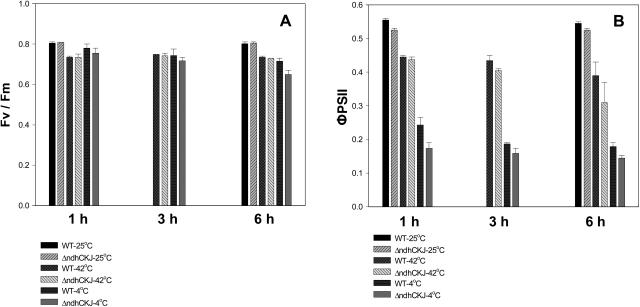
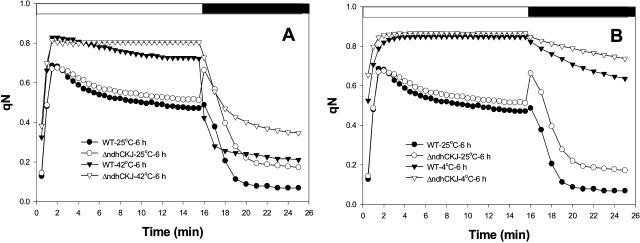
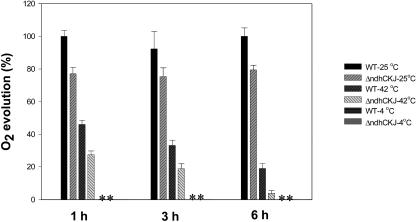
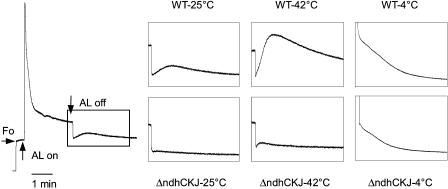
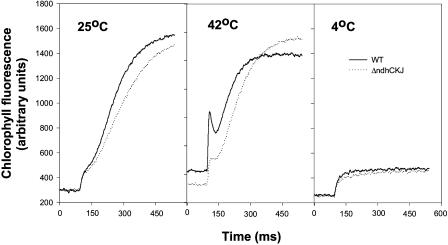
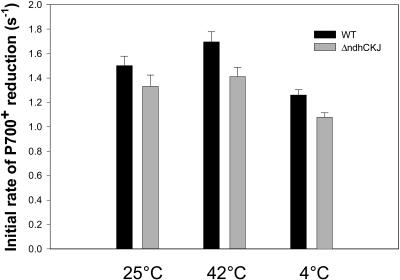
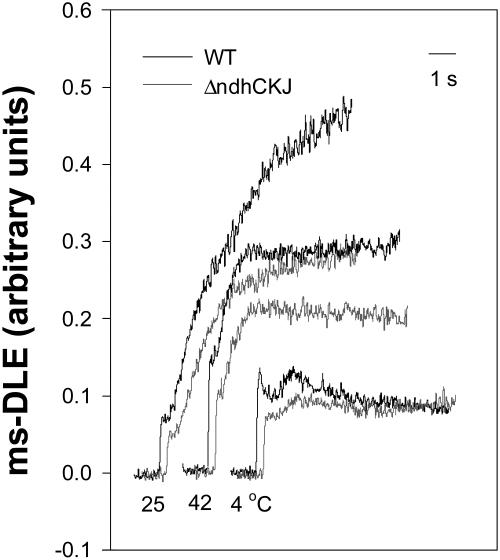
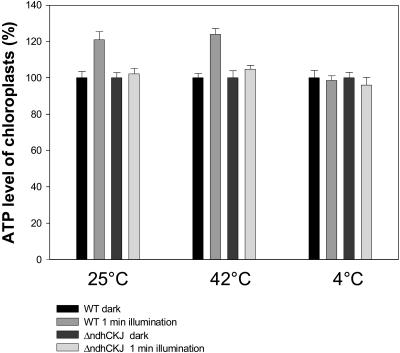
In this study, the function of the NAD(P)H dehydrogenase (NDH)-dependent pathway in suppressing the accumulation of reactive oxygen species in chloroplasts was investigated. Hydrogen peroxide accumulated in the leaves of tobacco (Nicotiana tabacum) defective in ndhC-ndhK-ndhJ (DeltandhCKJ) at 42 degrees C and 4 degrees C, and in that of wild-type leaves at 4 degrees C. The maximum quantum efficiency of PSII decreased to a similar extent in both strains at 42 degrees C, while it decreased more evidently in DeltandhCKJ at 4 degrees C. The parameters linked to CO(2) assimilation, such as the photochemical efficiency of PSII, the decrease of nonphotochemical quenching following the initial rise, and the photosynthetic O(2) evolution, were inhibited more significantly in DeltandhCKJ than in wild type at 42 degrees C and were seriously inhibited in both strains at 4 degrees C. While cyclic electron flow around PSI mediated by NDH was remarkably enhanced at 42 degrees C and suppressed at 4 degrees C. The proton gradient across the thylakoid membranes and light-dependent ATP synthesis were higher in wild type than in DeltandhCKJ at either 25 degrees C or 42 degrees C, but were barely formed at 4 degrees C. Based on these results, we suggest that cyclic photophosphorylation via the NDH pathway might play an important role in regulation of CO(2) assimilation under heat-stressed condition but is less important under chilling-stressed condition, thus optimizing the photosynthetic electron transport and reducing the generation of reactive oxygen species.
Figures
References
-
- Allen DJ, Ort DR (2001) Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci 6: 36–42 - PubMed
-
- Arnon DI, Allen MB, Whatley FR (1954) Photosynthesis by isolated chloroplasts. Nature 174: 394–396 - PubMed
-
- Asada K (1992) Ascorbate peroxidase: a hydrogen peroxide scavenging enzyme in plants. Physiol Plant 85: 235–241
-
- Asada K (1999) The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol Biol 50: 601–639 - PubMed
-
- Asada K, Kiso K (1973) Initiation of aerobic oxidation of sulfite by illuminated spinach chloroplasts. Eur J Biochem 33: 253–257 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
