

The interaction between DCL1 and HYL1 is important for efficient and precise processing of pri-miRNA in plant microRNA biogenesis
- PMID: 16428603
- PMCID: PMC1370900
- DOI: 10.1261/rna.2146906
The interaction between DCL1 and HYL1 is important for efficient and precise processing of pri-miRNA in plant microRNA biogenesis
Abstract
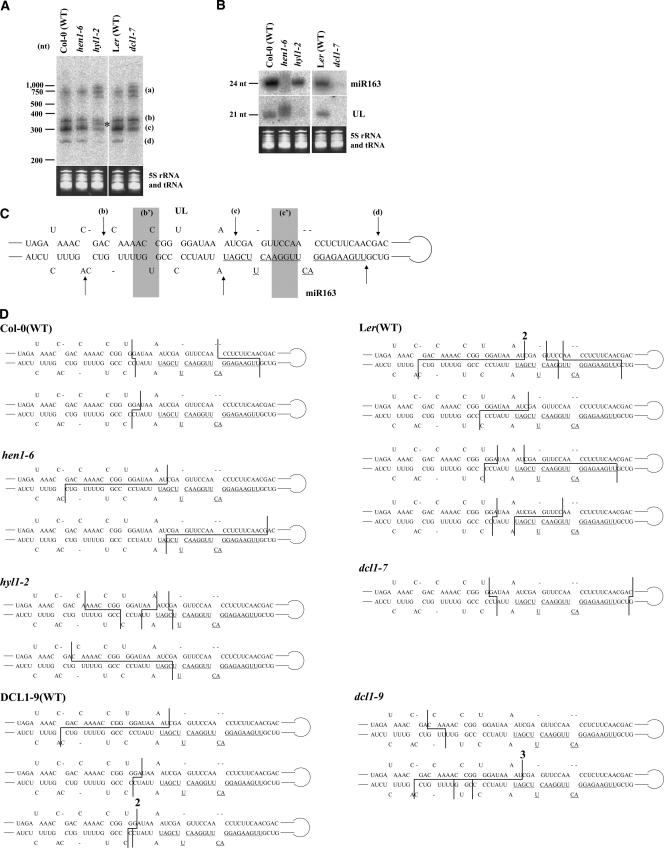
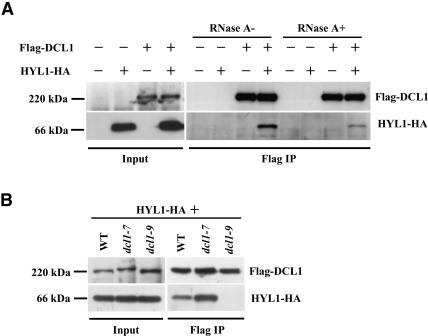
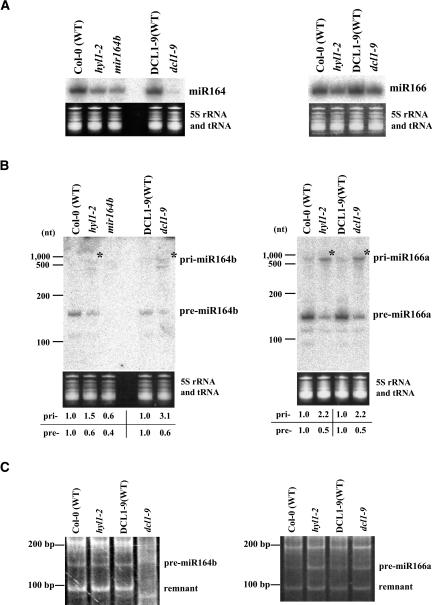
It has been reported that some double-stranded RNA (dsRNA) binding proteins interact with small RNA biogenesis-related RNase III enzymes. However, their biological significance is poorly understood. Here we examine the relationship between the Arabidopsis microRNA- (miRNA) producing enzyme DCL1 and the dsRNA binding protein HYL1. In the hyl1-2 mutant, the processing steps of miR163 biogenesis were partially impaired; increased accumulation of pri-miR163 and reduced accumulation of short pre-miR163 and mature miR163 as well as misplaced cleavages in the stem structure of pri-miR163 were detected. These misplaced cleavages were similar to those previously observed in the dcl1-9 mutant, in which the second double-stranded RNA binding domain of the protein was disrupted. An immunoprecipitation assay using Agrobacterium-mediated transient expression in Nicotiana benthamiana showed that HYL1 was able to form a complex with wild-type DCL1 protein, but not with the dcl1-9 mutant protein. We also examined miR164b and miR166a biogenesis in hyl1-2 and dcl1-9. Increased accumulation of pri-miRNAs and reduced accumulation of pre-miRNAs and mature miRNAs were detected. Misplaced cleavage on pri-miR164b was observed only in dcl1-9 but not in hyl1-2, whereas not on pri-miR166a in either mutant. These results indicate that HYL1 has a function in assisting efficient and precise cleavage of pri-miRNA through interaction with DCL1.
Figures
References
-
- Allen, E., Xie, Z., Gustafson, A.M., Sung, G.H., Spatafora, J.W., and Carrington, J.C. 2004. Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat. Genet. 12: 1282–1290. - PubMed
-
- Aoyama, T. and Chua, N.H. 1997. A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 11: 605– 612. - PubMed
-
- Denli, A.M., Tops, B.B.J., Plasterk, R.H.A., Ketting, R.F., and Hannon, G.J. 2004. Processing of primary microRNAs by the Microprocessor complex. Nature 432: 231–235. - PubMed
-
- Gregory, R.I., Yan, K.P., Amuthan, G., Chendrimada, T., Doratotaj, B., Cooch, N., and Shiekhattar, R. 2004. The Microprocessor complex mediates the genesis of microRNAs. Nature 432: 235–240. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases