

doi: 10.1128/IAI.74.2.1407-1411.2006.
Human tumor necrosis factor is a chemoattractant for the parasite Entamoeba histolytica
Affiliations
- PMID: 16428794
- PMCID: PMC1360330
- DOI: 10.1128/IAI.74.2.1407-1411.2006
Item in Clipboard
Human tumor necrosis factor is a chemoattractant for the parasite Entamoeba histolytica
Infect Immun.
2006 Feb.
Abstract
In an analysis of the molecular factors triggering amebiasis, we investigated the chemotaxis of Entamoeba histolytica toward tumor necrosis factor (TNF) in vitro, using quantitative imaging techniques. Our findings enabled us to propose a hitherto unknown role for TNF as a chemokinetic and chemoattractant agent for this parasite.
Figures
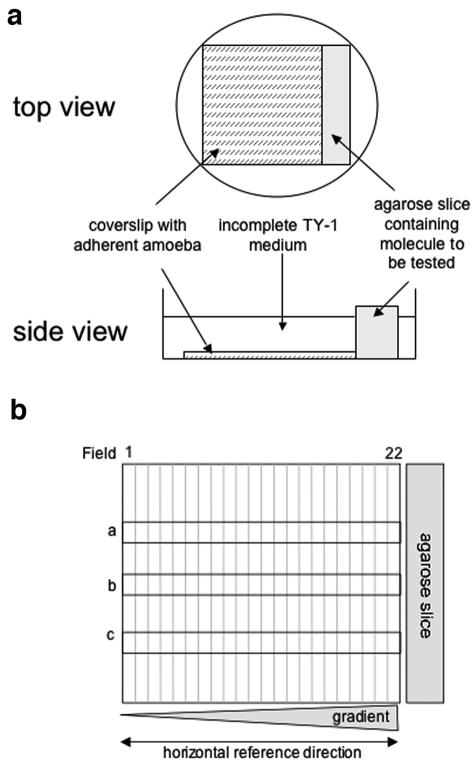
“Chemotaxis-on-coverslip assay” setup. (a) Experimental setup. One milliliter of incomplete TY-1 medium, without decomplemented serum, vitamins, or penicillin-streptomycin (iTY-1), containing 2.5 × 105 E. histolytica trophozoites previously treated with fluorescent CellTracker green (Molecular Probes) was deposited onto a coverslip placed in a 35-mm culture dish. The trophozoites were allowed to adhere for 10 min at 37°C, during which time the test molecules were injected into a slice of 1% agarose (20 mm by 5 mm by 5 mm). The excess medium was removed from the dishes, the agarose slice was placed next to the coverslip, and 1.5 ml of iTY-1 was then added to the dish. E. histolytica is an anaerobic organism, so the level of oxygen was minimized by placing the dishes in an anaerobic environment (GENbag anaer; Biomerieux). The cells were incubated at 37°C for 2 hours. The trophozoites were fixed with 3.7% paraformaldehyde for 20 min at room temperature. The coverslips were mounted onto glass slides so that the cell distribution could be examined. The progression of Ponceau red (5 μl of a 3.75% solution injected into the agarose slice) was monitored as a measure of the stability of the gradient. A stable, linear gradient was found at between 1 and 3 h. (b) To quantify the cell distribution, the trophozoites were imaged with a Zeiss inverted microscope in epifluorescence mode (fluorescein filters), running the “mosaic” option of the Simple PCI software. The coverslip's vertical axis was divided into 22 fields, with field 22 nearest the agarose slice. Three horizontal rows (a, b, and c) of 22 adjacent images were taken along the coverslip (covering 10% of the coverslip's area). The number of trophozoites was counted in each field for rows a, b, and c, so as to obtain the total number of amebae per field. The total population counted was obtained by summing the number of parasites in rows a, b, and c for the 22 fields.
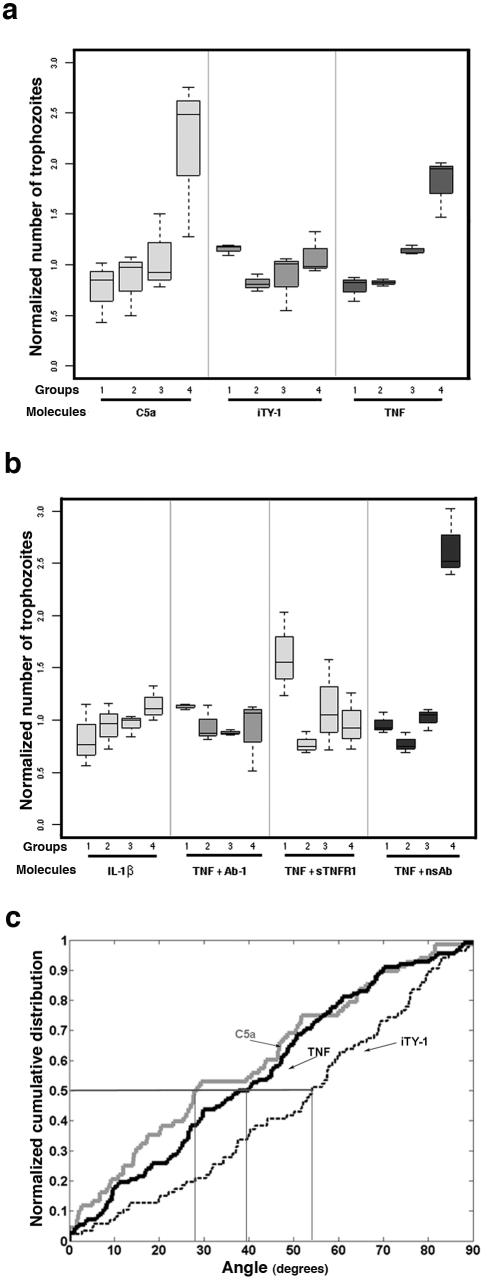
TNF has a chemoattractant effect on E. histolytica. The displacement of the E. histolytica population was analyzed after 2 h of incubation in the presence of different molecules. The box plot shows the distribution of cells along the coverslip in groups 1 (fields 1 to 6), 2 (fields 7 to 11), 3 (fields 12 to 17), and 4 (fields 18 to 22). A one-way analysis of variance allowed us to compare the difference in amebae numbers between groups. (a) In the absence of a chemoattractant (agarose with iTY-1 [without serum]), the population was distributed homogeneously along the coverslip (P = 0.8). When the agarose slice was filled with C5a (6 nM) or TNF (50 nM), there was a displacement of the population up the gradient toward the test molecule source (C5a, P = 8.7 × 10−3; TNF, P = 1.6 × 10−4). In addition, a pairwise comparison using t tests between groups was applied. In the C5a gradient, the mean ameba number was significantly different between groups 1 and 4 (P = 0.008), groups 2 and 4 (P = 0.011), and groups 3 and 4 (P = 0.025), with an accumulation of amebae near the source of C5a. The mean per group increased along the gradient of TNF and was significantly different between groups 1 and 3 (P = 0.025), groups 1 and 4 (P = 0.015), groups 2 and 3 (P < 10−4), and groups 2 and 4 (P = 0.03). In contrast, for iTY-1 no significant difference between groups was found, in accord with an absence of chemotactic migration. (b) In the presence of a gradient of IL-1β (source, 50 nM) there was no displacement of the population (P = 0.08). When TNF (50 nM; 0.88 μg/ml) was incubated with a monoclonal anti-TNF antibody (500 μg/ml) (TNF + Ab-1) or with the soluble TNF receptor I (4 μg/ml) (TNF + sTNFRI) prior to injection into the agarose cube, chemotaxis was abrogated (P = 0.14 and P = 0.16, respectively). However, when TNF was incubated with a monoclonal antibody (500 μg/ml) not specific for human TNF (TNF + nsAb), chemotaxis toward the TNF source was not inhibited (P = 0.003). Total numbers of ameba analyzed: iTY-1, n = 3,303; IL-1β, n = 4,022; C5a, n = 8,385; TNF, n = 7,871; TNF + Ab-1, n = 7,527; TNF + sTNFRI, n = 1,677; TNF + nsAb, n = 1,927. (c) Normalized cumulative distribution of cell orientations. The distributions of orientations of cells moving toward C5a (6 nM) and TNF (50 nM) were significantly different from the distribution in iTY-1 (P < 0.0006 for C5a and P < 0.0025 for TNF). In absence of chemotaxis (iTY-1), the median angle value was 54.0°. In presence of a C5a or TNF gradient, the median angle values were 28.2° and 38.8°, respectively, showing that the majority of cells oriented preferentially along the gradient, as expected for chemotaxis. Total numbers of amebae analyzed: iTY-1, n = 86; C5a, n = 68; TNF, n = 112.
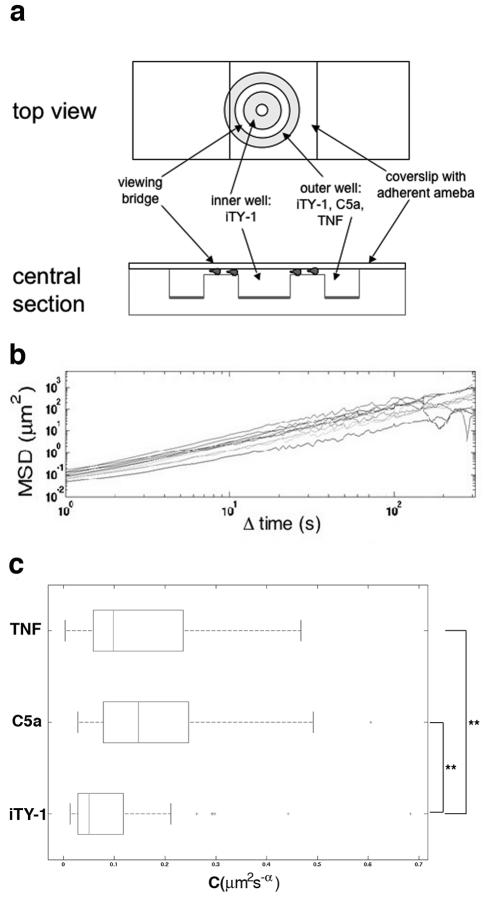
TNF has a chemokinetic effect on E. histolytica. (a) Dunn chemotaxis chamber (20). The two circular wells were filled with iTY-1. A suspension of amebae was placed on a coverslip, and amebae were allowed to adhere for 10 min at 37°C. The coverslip was turned over and placed on the chamber so that the amebae bathed in the iTY-1. Once the chamber was set up, the outer well was emptied using a syringe and refilled with iTY-1 or the test molecule, C5a (6 nM) or TNF (30 nM). Phase-contrast images of moving cells were recorded by video microscopy at 1-second intervals for 45 min under a Zeiss inverted light microscope using a 10× objective in transmission mode. (b) Mean square displacements for 10 individual cells (different lines), calculated from the parasite trajectories obtained by computerized tracking (21) of the 300 images from 25 to 30 min. The approximate linearity of these curves on a logarithmic scale means that the mean square displacements fit well to power laws 〈(Δr2)〉 = C(Δt)α. The roughly constant exponents α ≈ 1.5 were equivalent to those found for E. histolytica in the absence of chemotactic gradients (6), thus indicating that E. histolytica chemotactic motility is superdiffusive. The constancy of the exponent enables us to use the coefficient C to measure each cell's rate of movement. (c) Rate of movement of E. histolytica during chemotaxis. Box plots of C for cells in iTY-1, C5a, and TNF (n = 58) are shown. E. histolytica trophozoites present in C5a and TNF gradients have a significantly higher rate of movement than trophozoites in iTY-1 (3 and 2 times, respectively). **, significantly different, with α < 0.01. Total numbers of ameba analyzed: iTY-1, n = 53; C5a, n = 43; TNF, n = 58.
References
-
- Acevado, J. A., J. Pacheco-Yépez, J. Serrano-Luna, M. Espinosa-Cantallano, V. Tsutsumi, and M. Shibayama. 2000. Experimental amebic liver abscess: in vivo localization of TNF-α. Arch. Med. Res. 31:S98-S100. - PubMed
-
- Alon, R., L. Cahalon, R. Hershkoviz, D. Elbaz, B. Reizis, D. Wallach, S. K. Akiyama, K. M. Yamada, and O. Lider. 1994. TNF-alpha binds to the N-terminal domain of fibronectin and augments the beta 1-integrin-mediated adhesion of CD4+ T lymphocytes to the glycoprotein. J. Immunol. 152:1304-1313. - PubMed
-
- Bailey, G. B., G. J. Leitch, and D. B. Day. 1985. Chemotaxis by Entamoeba histolytica. J. Protozool. 32:341-346. - PubMed
-
- Chen, W. N., R. L. Woodbury, L. E. Kathmann, L. K. Opresko, R. C. Zangar, H. S. Wiley, and B. D. Thrall. 2004. Induced autocrine signaling through the epidermal growth factor receptor contributes to the response of mammary epithelial cells to tumor necrosis factor alpha. J. Biol. Chem. 279:18488-18496. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
