

The Runx1/AML1 transcription factor selectively regulates development and survival of TrkA nociceptive sensory neurons
- PMID: 16429136
- PMCID: PMC2703717
- DOI: 10.1038/nn1631
The Runx1/AML1 transcription factor selectively regulates development and survival of TrkA nociceptive sensory neurons
Abstract
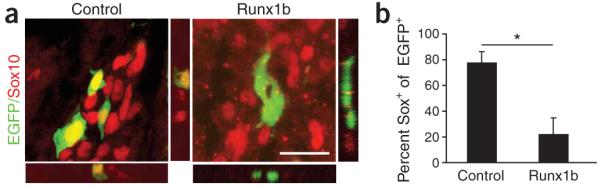
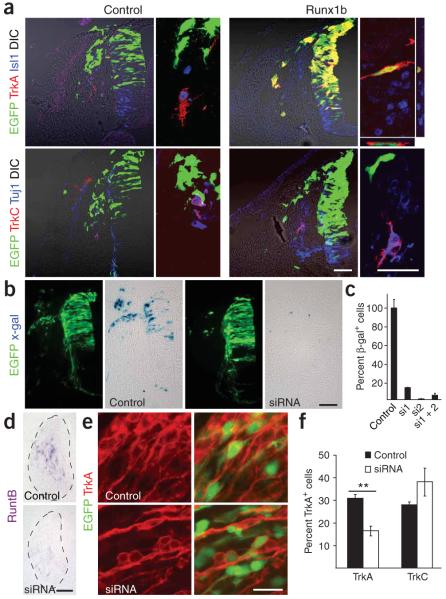
Neural crest cells (NCCs) can adopt different neuronal fates. In NCCs, neurogenin-2 promotes sensory specification but does not specify different subclasses of sensory neurons. Understanding the gene cascades that direct Trk gene activation may reveal mechanisms generating sensory diversity, because different Trks are expressed in different sensory neuron subpopulations. Here we show in chick and mouse that the Runt transcription factor Runx1 promotes axonal growth, is selectively expressed in neural crest-derived TrkA(+) sensory neurons and mediates TrkA transactivation in migratory NCCs. Inhibition of Runt activity depletes TrkA expression and leads to neuronal death. Moreover, Runx1 overexpression is incompatible with multipotency in the migratory neural crest but does not induce expression of pan-neuronal genes. Instead, Runx1-induced neuronal differentiation depends on an existing neurogenin2 proneural gene program. Our data show that Runx1 directs, in a context-dependent manner, key aspects of the establishment of the TrkA(+) nociceptive subclass of neurons.
Figures
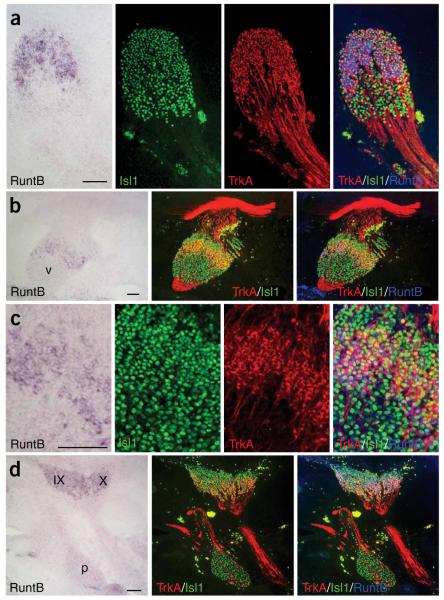
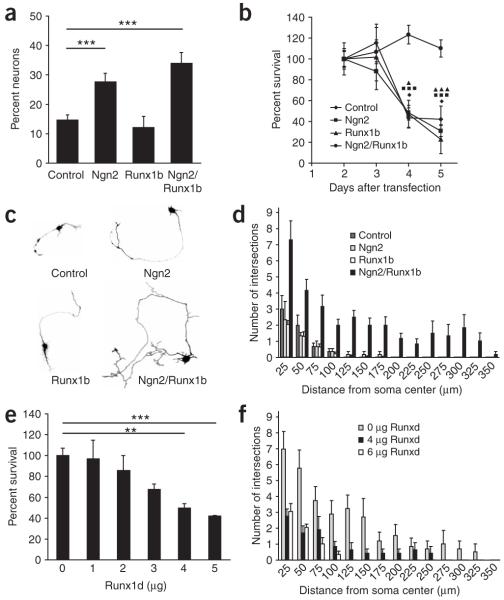
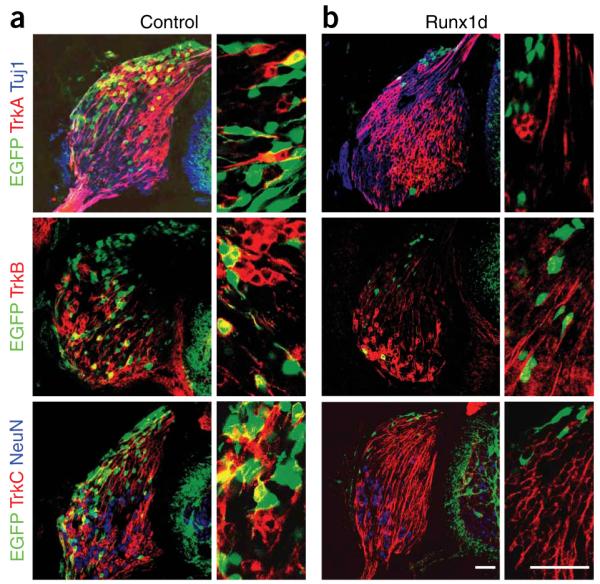
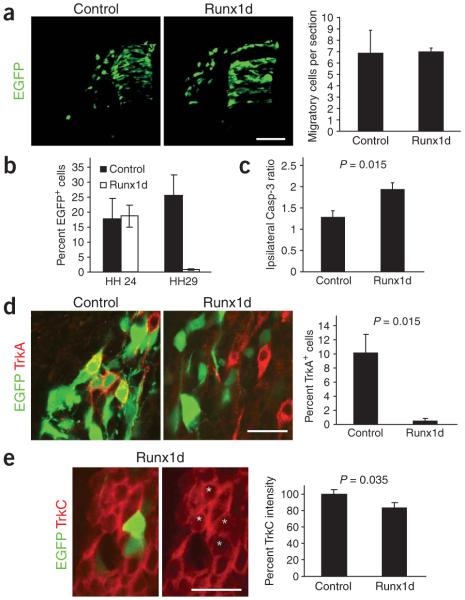
Comment in
-
Specifying nociceptors.Nat Neurosci. 2006 Feb;9(2):162. doi: 10.1038/nn0206-162. Nat Neurosci. 2006. PMID: 16439982 No abstract available.
References
-
- Carr PA, Nagy JI. Emerging relationships between cytochemical properties and sensory modality transmission in primary sensory neurons. Brain Res. Bull. 1993;30:209–219. - PubMed
-
- Cheung M, et al. The transcriptional control of trunk neural crest induction, survival, and delamination. Dev. Cell. 2005;8:179–192. - PubMed
-
- Kim J, Lo L, Dormand E, Anderson DJ. SOX10 maintains multipotency and inhibits neuronal differentiation of neural crest stem cells. Neuron. 2003;38:17–31. - PubMed
-
- Serbedzija GN, Fraser SE, Bronner-Fraser M. Pathways of trunk neural crest cell migration in the mouse embryo as revealed by vital dye labelling. Development. 1990;108:605–612. - PubMed
-
- Kasemeier-Kulesa JC, Kulesa PM, Lefcort F. Imaging neural crest cell dynamics during formation of dorsal root ganglia and sympathetic ganglia. Development. 2005;132:235–245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
