

RNA nanotechnology: engineering, assembly and applications in detection, gene delivery and therapy
- PMID: 16430131
- PMCID: PMC2842999
- DOI: 10.1166/jnn.2005.446
RNA nanotechnology: engineering, assembly and applications in detection, gene delivery and therapy
Abstract
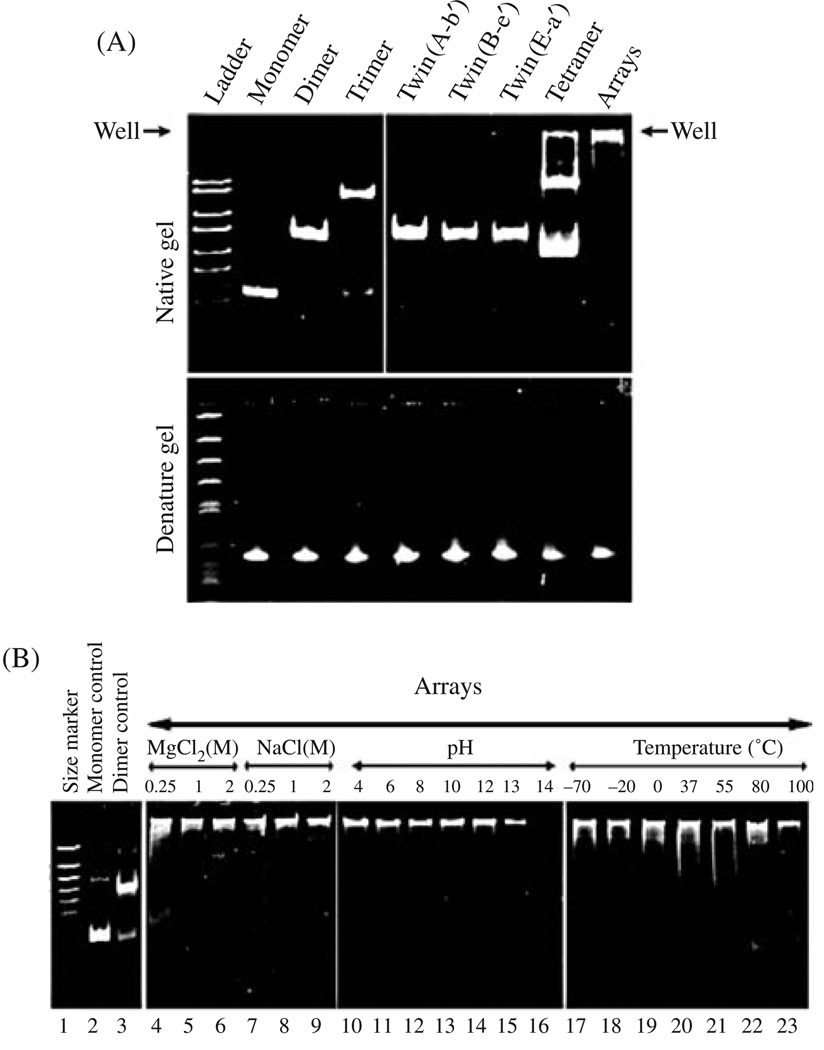
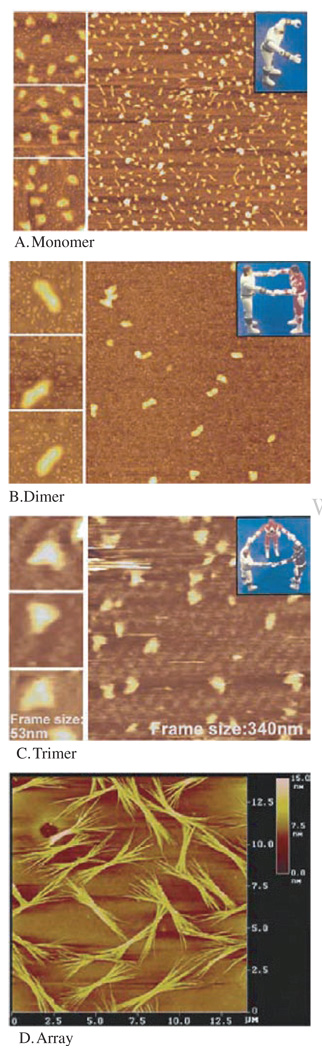
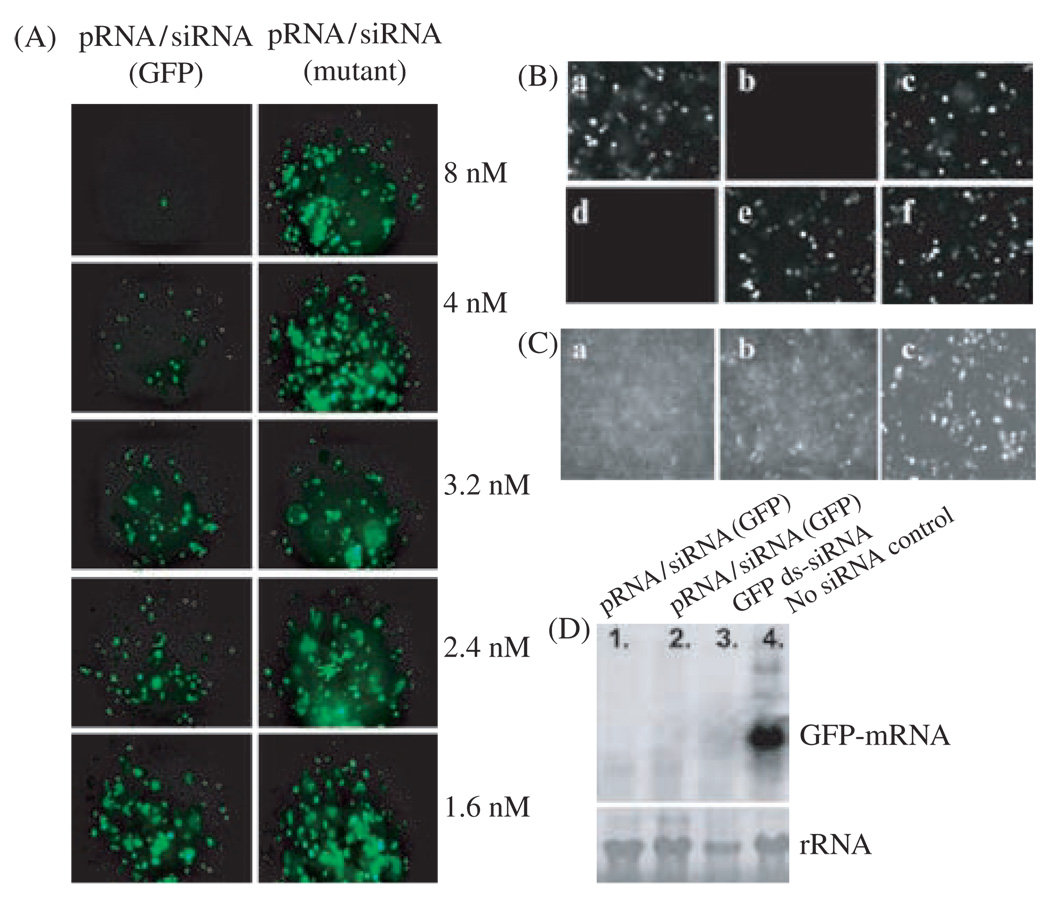
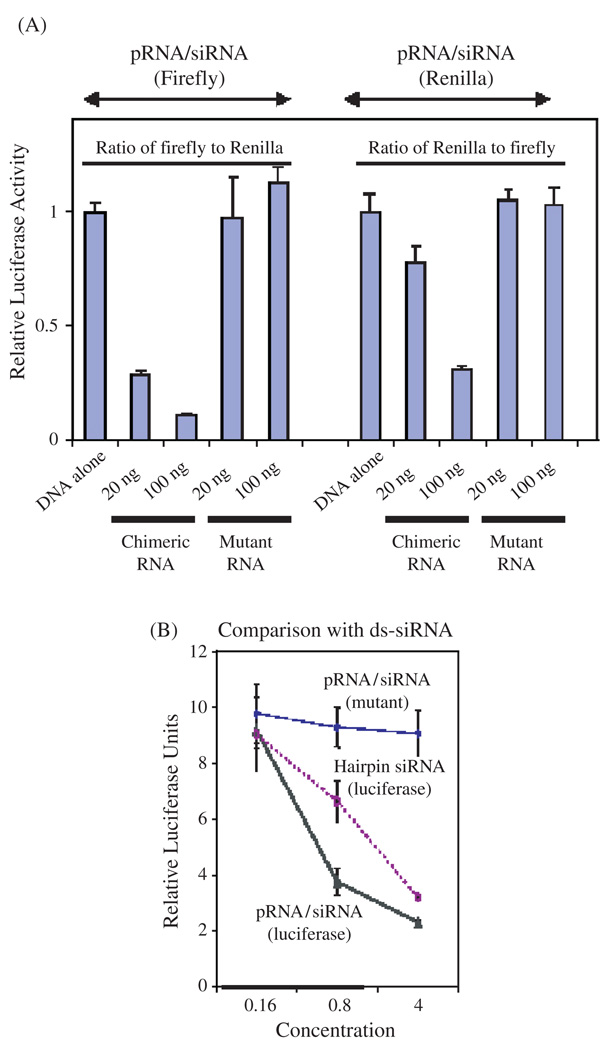
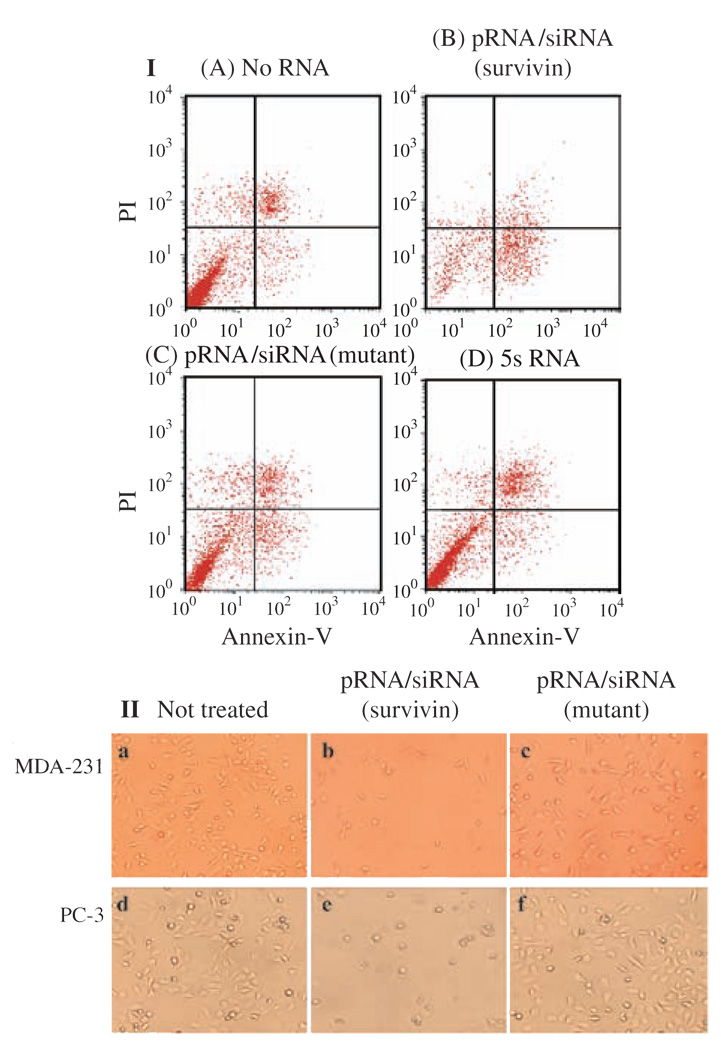
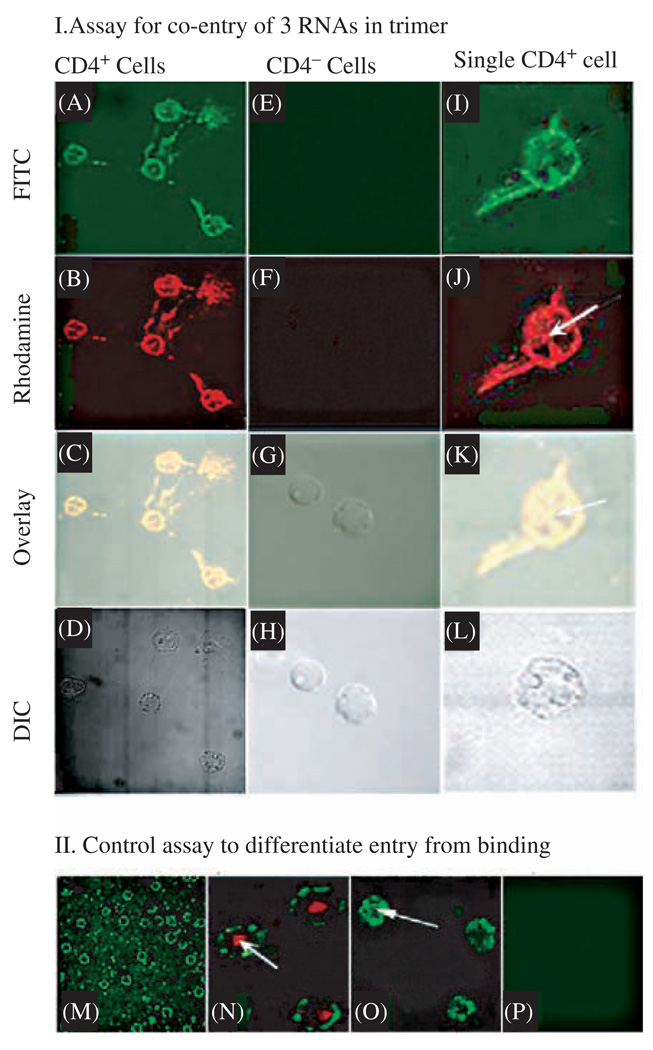
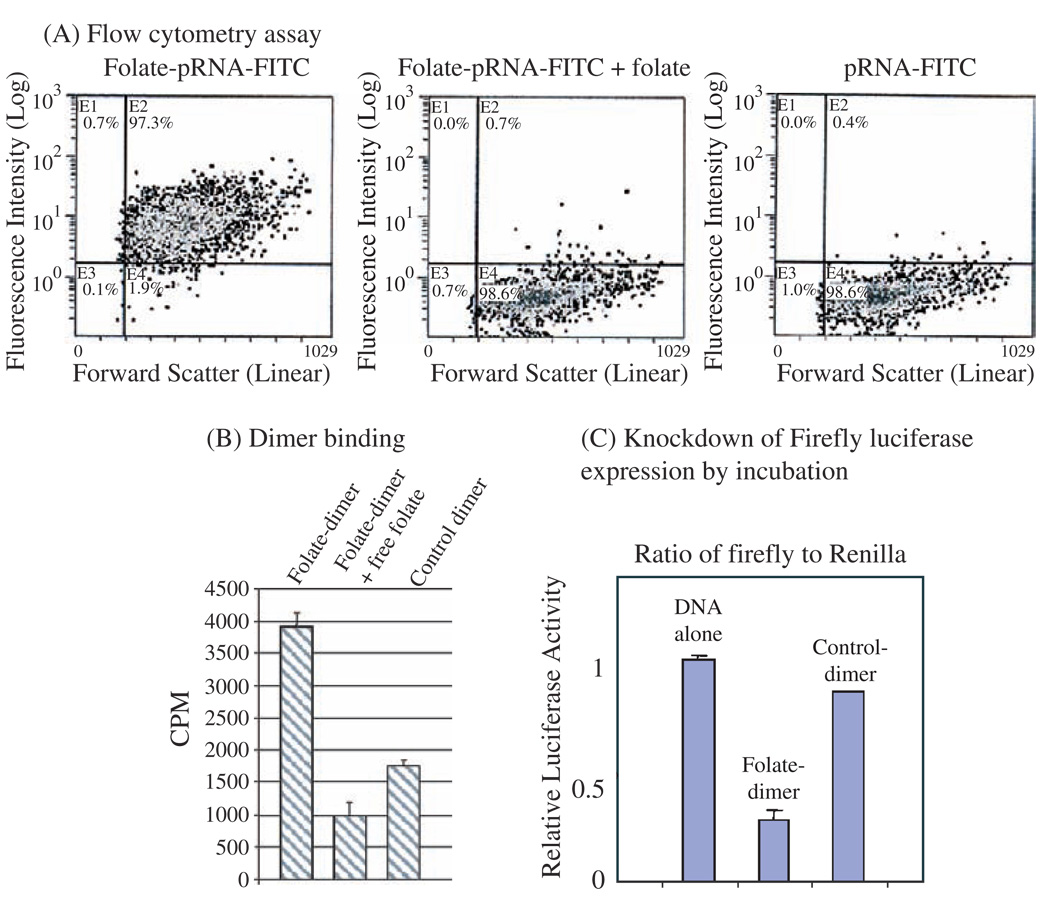
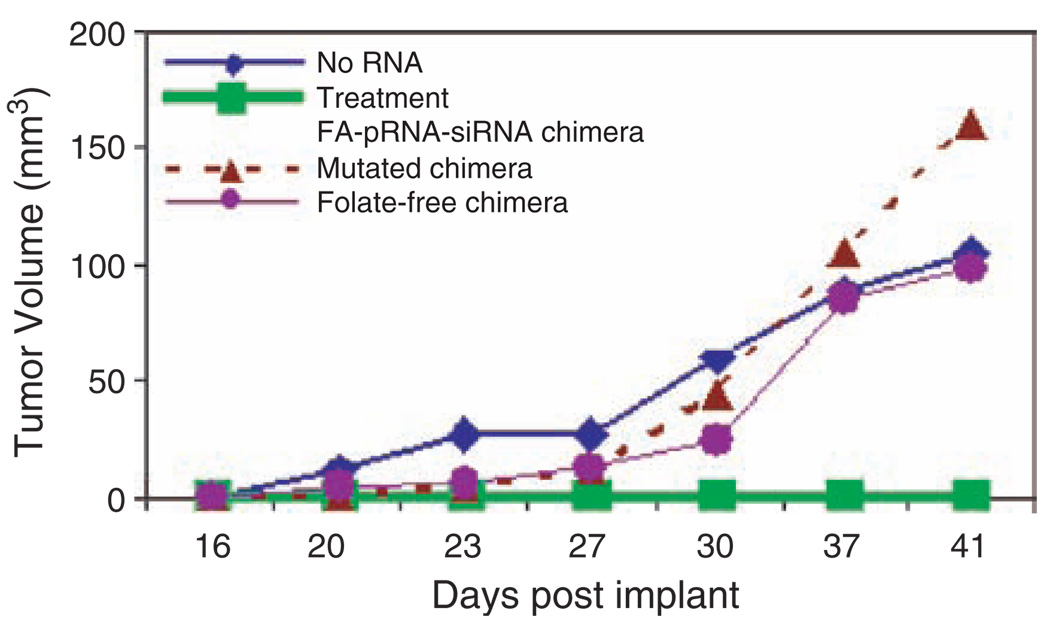
Biological macromolecules including DNA, RNA, and proteins, have intrinsic features that make them potential building blocks for the bottom-up fabrication of nanodevices. RNA is unique in nanoscale fabrication due to its amazing diversity of function and structure. RNA molecules can be designed and manipulated with a level of simplicity characteristic of DNA while possessing versatility in structure and function similar to that of proteins. RNA molecules typically contain a large variety of single stranded loops suitable for inter- and intra-molecular interaction. These loops can serve as mounting dovetails obviating the need for external linking dowels in fabrication and assembly. The self-assembly of nanoparticles from RNA involves cooperative interaction of individual RNA molecules that spontaneously assemble in a predefined manner to form a larger two- or three-dimensional structure. Within the realm of self-assembly there are two main categories, namely template and non-template. Template assembly involves interaction of RNA molecules under the influence of specific external sequence, forces, or spatial constraints such as RNA transcription, hybridization, replication, annealing, molding, or replicas. In contrast, non-template assembly involves formation of a larger structure by individual components without the influence of external forces. Examples of non-template assembly are ligation, chemical conjugation, covalent linkage, and loop/loop interaction of RNA, especially the formation of RNA multimeric complexes. The best characterized RNA multiplier and the first to be described in RNA nanotechnological application is the motor pRNA of bacteriophage phi29 which form dimers, trimers, and hexamers, via hand-in-hand interaction. phi29 pRNA can be redesigned to form a variety of structures and shapes including twins, tetramers, rods, triangles, and 3D arrays several microns in size via interaction of programmed helical regions and loops. 3D RNA array formation requires a defined nucleotide number for twisting and a palindromic sequence. Such arrays are unusually stable and resistant to a wide range of temperatures, salt concentrations, and pH. Both the therapeutic siRNA or ribozyme and a receptor-binding RNA aptamer or other ligands have been engineered into individual pRNAs. Individual chimeric RNA building blocks harboring siRNA or other therapeutic molecules have been fabricated subsequently into a trimer through hand-in-hand interaction of the engineered right and left interlocking RNA loops. The incubation of these particles containing the receptor-binding aptamer or other ligands results in the binding and co-entry of trivalent therapeutic particles into cells. Such particles were subsequently shown to modulate the apoptosis of cancer cells in both cell cultures and animal trials. The use of such antigen-free 20-40 nm particles holds promise for the repeated long-term treatment of chronic diseases. Other potentially useful RNA molecules that form multimers include HIV RNA that contain kissing loop to form dimers, tecto-RNA that forms a "jigsaw puzzle," and the Drosophila bicoid mRNA that forms multimers via "hand-by-arm" interactions. Applications of RNA molecules involving replication, molding, embossing, and other related techniques, have recently been described that allow the utilization of a variety of materials to enhance diversity and resolution of nanomaterials. It should eventually be possible to adapt RNA to facilitate construction of ordered, patterned, or pre-programmed arrays or superstructures. Given the potential for 3D fabrication, the chance to produce reversible self-assembly, and the ability of self-repair, editing and replication, RNA self-assembly will play an increasingly significant role in integrated biological nanofabrication. A random 100-nucleotide RNA library may exist in 1.6 x 10(60) varieties with multifarious structure to serve as a vital system for efficient fabrication, with a complexity and diversity far exceeding that of any current nanoscale system. This review covers the basic concepts of RNA structure and function, certain methods for the study of RNA structure, the approaches for engineering or fabricating RNA into nanoparticles or arrays, and special features of RNA molecules that form multimers. The most recent development in exploration of RNA nanoparticles for pathogen detection, drug/gene delivery, and therapeutic application is also introduced in this review.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical