

A plausible model of phyllotaxis
- PMID: 16432192
- PMCID: PMC1345713
- DOI: 10.1073/pnas.0510457103
A plausible model of phyllotaxis
Abstract
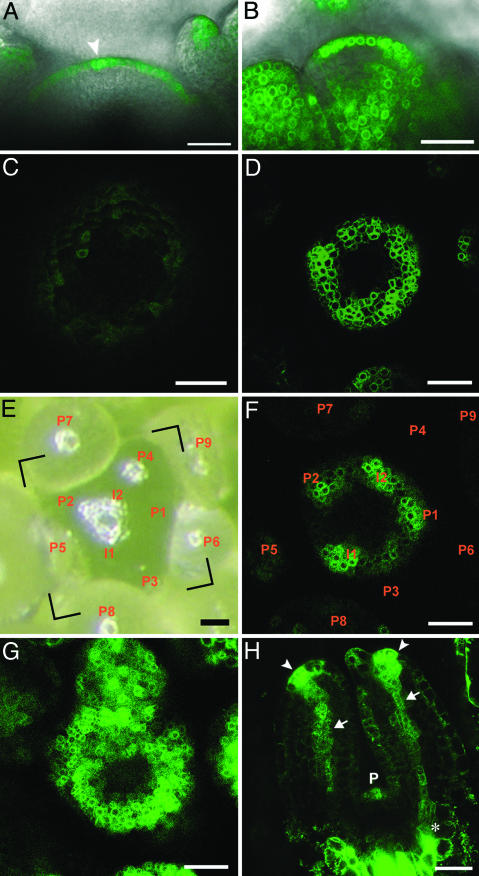
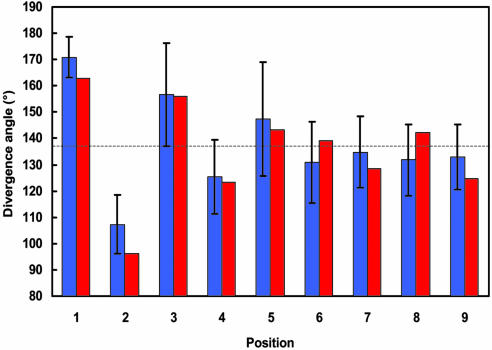
A striking phenomenon unique to the kingdom of plants is the regular arrangement of lateral organs around a central axis, known as phyllotaxis. Recent molecular-genetic experiments indicate that active transport of the plant hormone auxin is the key process regulating phyllotaxis. A conceptual model based on these experiments, introduced by Reinhardt et al. [Reinhardt, D., Pesce, E. R., Stieger, P., Mandel, T., Baltensperger, K., et al. (2003) Nature 426, 255-260], provides an intuitively plausible interpretation of the data, but raises questions of whether the proposed mechanism is, in fact, capable of producing the observed temporal and spatial patterns, is robust, can start de novo, and can account for phyllotactic transitions, such as the frequently observed transition from decussate to spiral phyllotaxis. To answer these questions, we created a computer simulation model based on data described previously or in this paper and reasonable hypotheses. The model reproduces, within the standard error, the divergence angles measured in Arabidopsis seedlings and the effects of selected experimental manipulations. It also reproduces distichous, decussate, and tricussate patterns. The model thus offers a plausible link between molecular mechanisms of morphogenesis and the geometry of phyllotaxis.
Figures



References
-
- Steeves, T. A. & Sussex, I. M. (1989) Patterns in Plant Development (Cambridge Univ. Press, Cambridge, U.K.).
-
- Hofmeister, W. (1868) in Handbuch der Physiologischen Botanik, eds. Bary, A. d., Irmisch, T. H. & Sachs, J. (Engelmann, Leipzig, Germany), pp. 405–664.
-
- Snow, M. & Snow, R. (1932) Philos. Trans. R. Soc. London B 221, 1–43.
-
- Richards, F. J. (1951) Philos. Trans. R. Soc. London B 235, 509–564.
-
- Green, P. B., Steele, C. S. & Rennich, S. C. (1996) Ann. Bot. (London) 77, 515–527.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
