

Involvement of the CA3-CA1 synapse in the acquisition of associative learning in behaving mice
- PMID: 16436593
- PMCID: PMC6674570
- DOI: 10.1523/JNEUROSCI.2834-05.2006
Involvement of the CA3-CA1 synapse in the acquisition of associative learning in behaving mice
Abstract
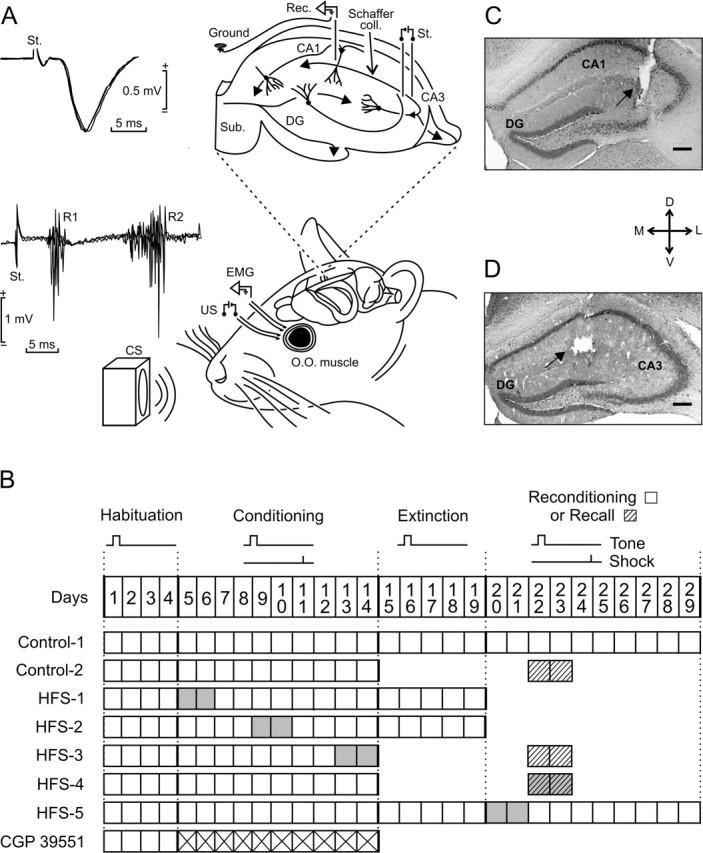
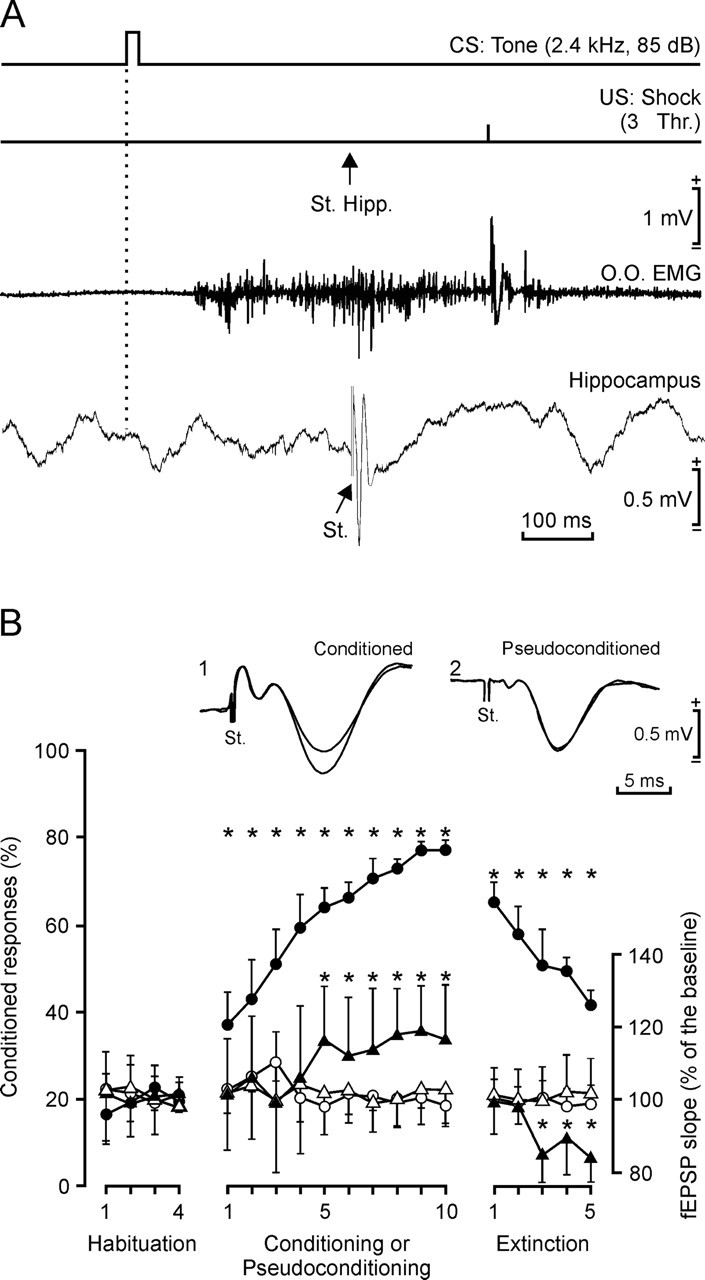
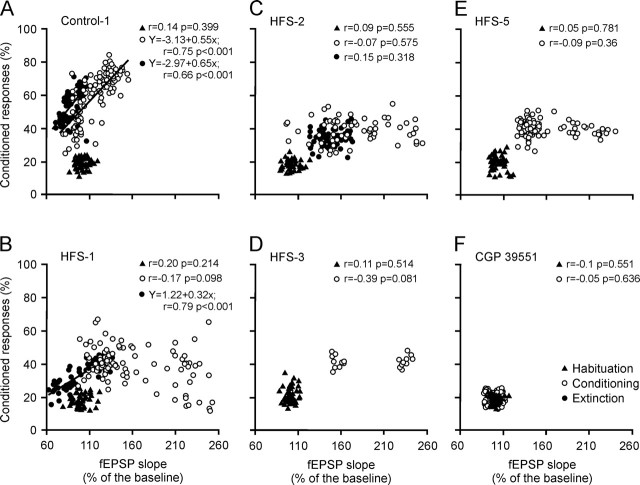
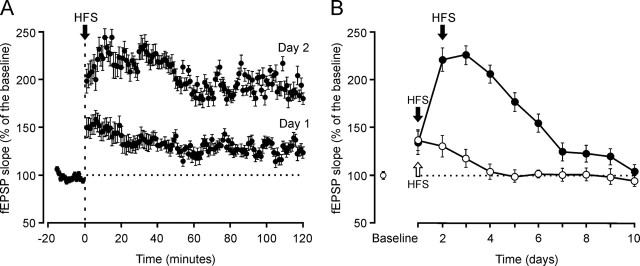
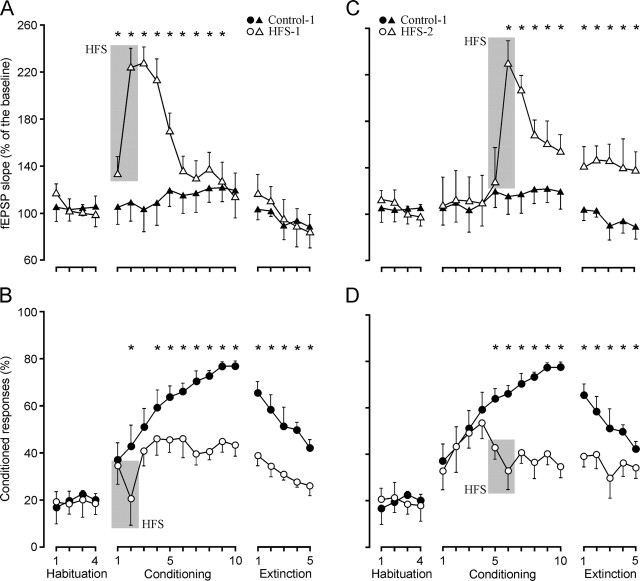
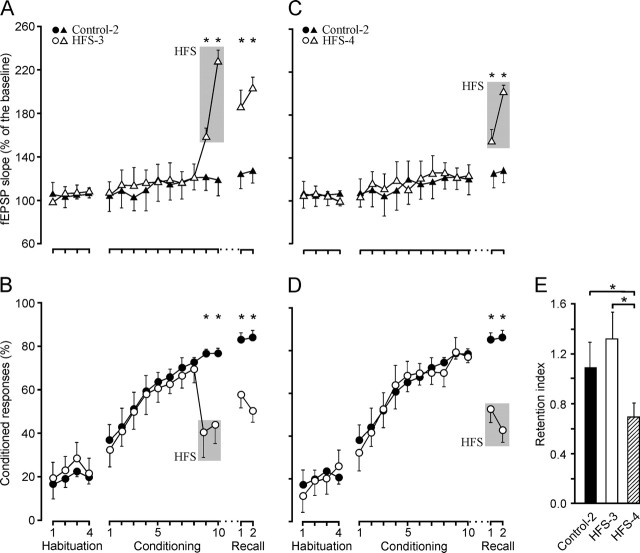
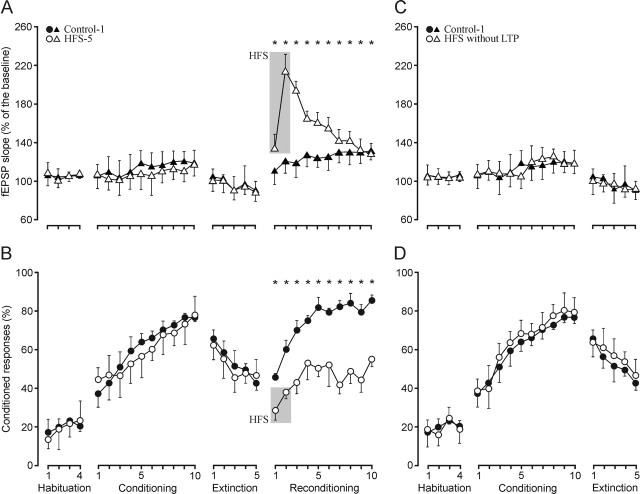
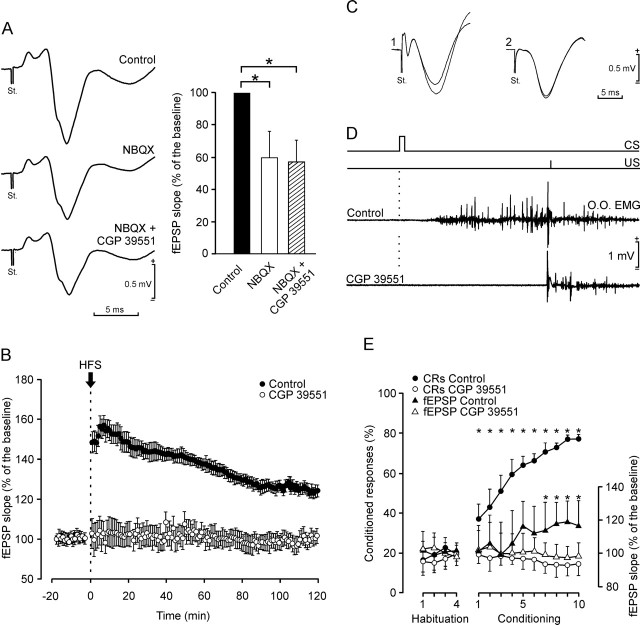
One of the brain sites more directly related with learning and memory processes is the hippocampus. We recorded, in conscious mice, the activity-dependent changes taking place at the hippocampal CA3-CA1 synapse during the acquisition, extinction, recall, and reconditioning of an associative task. Mice were classically conditioned to evoke eyelid responses using a trace [conditioned stimuli (CS), tone; unconditioned stimuli (US), shock] paradigm. A single electrical pulse presented to the Schaffer collateral-commissural pathway during the CS-US interval evoked a monosynaptic field EPSP (fEPSP) at ipsilateral CA1 pyramidal cells. The slope of evoked fEPSPs increased across conditioning sessions and decreased during extinction, being linearly related to learning evolution. In contrast, fEPSPs were not modified when evoked in control mice in the absence of a conditioning protocol. Long-term potentiation (LTP) evoked by high-frequency stimulation of Schaffer collaterals prevented acquisition, extinction, recall, or reconditioning, depending on the moment when it was triggered. Learning and memory impairments evoked by LTP induction resulted probably from the functional saturation of the CA3-CA1 synapse, although an additional disturbance of the subsequent information transfer toward postsynaptic circuits cannot be discarded. CGP 39551 [(E)-(+/-)-2-amino-4-methyl-5-phosphono-3-pentenoic acid ethyl ester] (an NMDA antagonist) prevented LTP induction in behaving mice, as well as the acquisition of an eyelid learned response, and the synaptic changes taking place at the CA3-CA1 synapse across conditioning. In conclusion, the responsivity of the CA3-CA1 synapse seems to be modulated during associative learning, and both processes are prevented by experimental LTP or NMDA-receptor inactivation. Our results provide evidence of a relationship between activity-dependent synaptic plasticity and associative learning in behaving mice.
Figures
References
-
- Berger TW, Rinaldi P, Weisz DJ, Thompson RF (1983). Single-unit analysis of different hippocampal cell types during classical conditioning of rabbit nictitating membrane response. J Neurophysiol 50:1197–1219. - PubMed
-
- Bliss TVP, Collingridge GL (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361:31–39. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous