

Dendrites contain a spacing pattern
- PMID: 16436602
- PMCID: PMC6674572
- DOI: 10.1523/JNEUROSCI.4424-05.2006
Dendrites contain a spacing pattern
Abstract
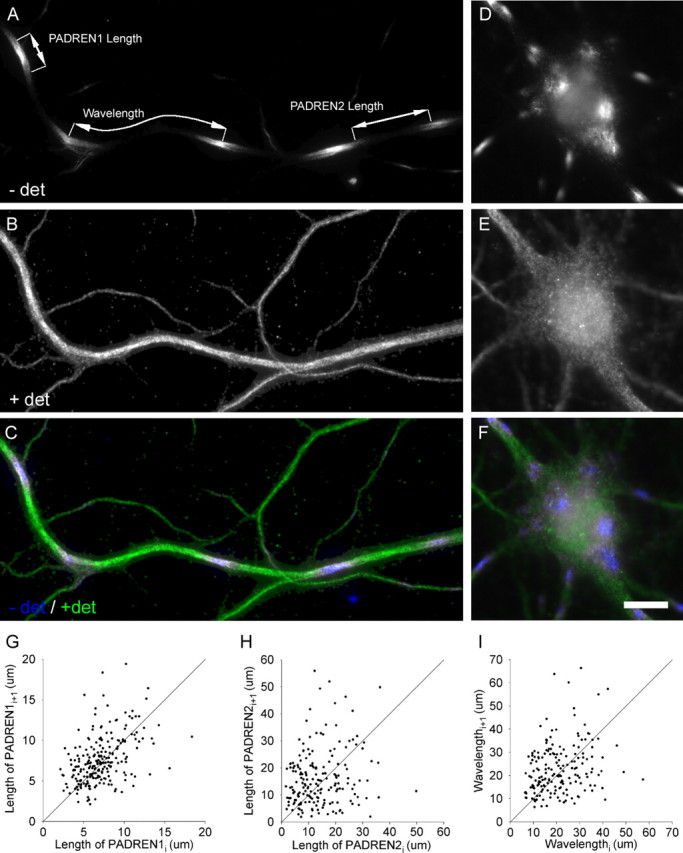
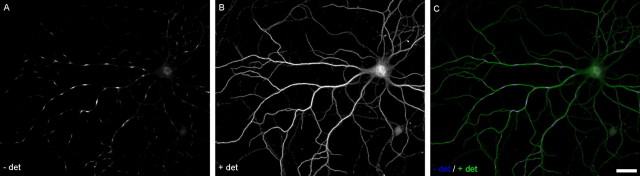
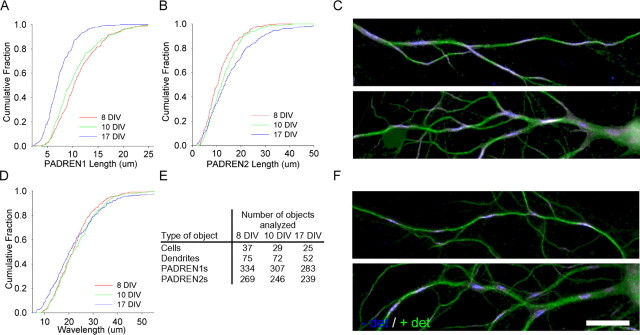
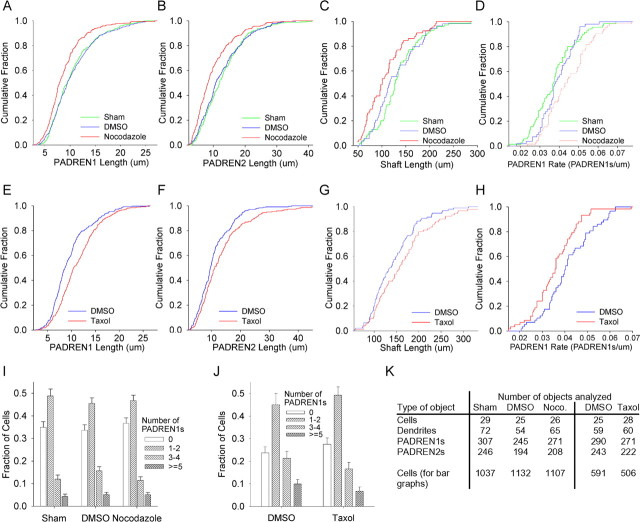
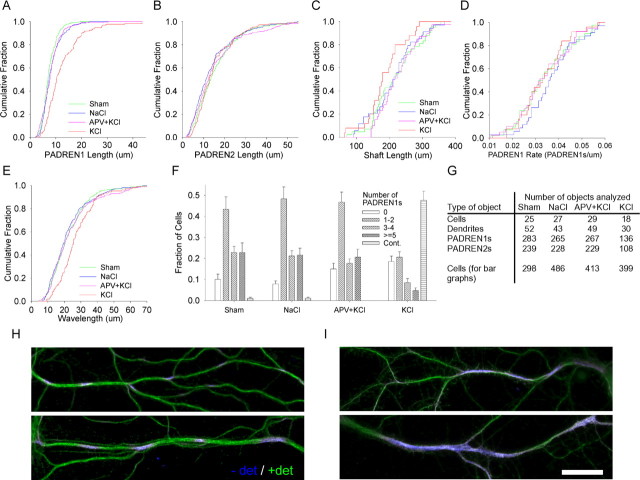
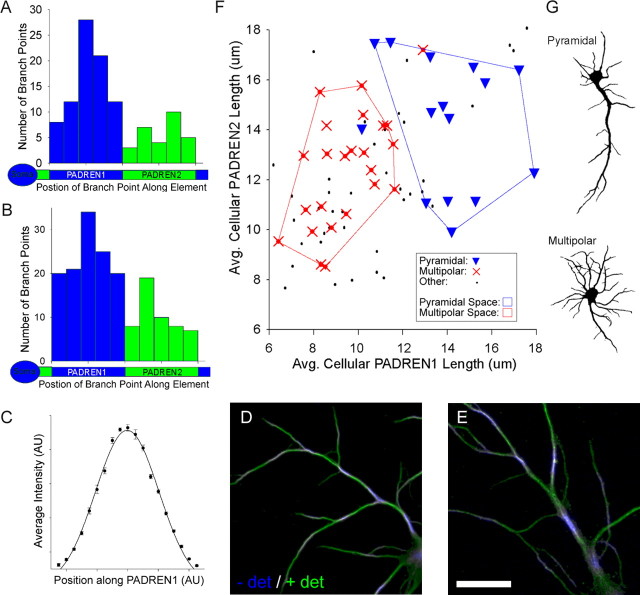
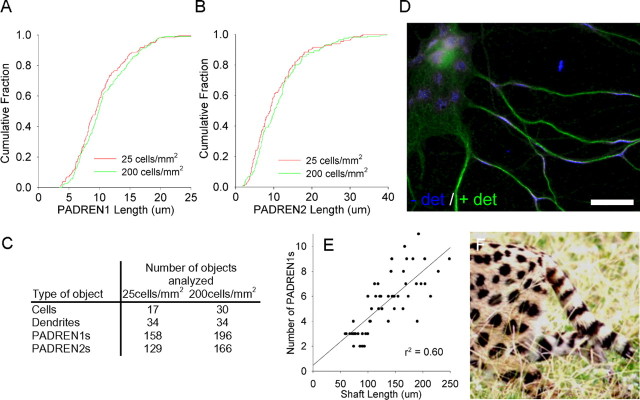
The distinctive branching patterns of dendritic arbors are essential for neuronal information processing. The final shape of an arbor is the result of intrinsic and extrinsic factors. However, the cellular mechanisms that underlie branch patterning are unknown. In many biological systems, locally acting factors are intrinsically organized into spacing patterns that guide patterned morphogenesis. Here, we show that neurons contain two types of periodic and regular elements (PADREN1s and PADREN2s) that are arranged into a spacing pattern. The wavelength of the pattern is approximately 20 microm. Dendritic branches occur preferentially within PADREN1s, and specific PADREN lengths correspond to specific arbor types. The lengths of the PADRENs also change over time and can be modified by activity. However, PADRENs are intrinsically organized, possibly by a reaction-diffusion process. PADRENs reveal a previously unrecognized level of neuronal organization that may provide insight into how the distinct branching patterns of the dendrites are intrinsically organized.
Figures
References
-
- Agarwala S, Sanders TA, Ragsdale CW (2001). Sonic hedgehog control of size and shape in midbrain pattern formation. Science 291:2147–2150. - PubMed
-
- Agmon-Snir H, Carr CE, Rinzel J (1998). The role of dendrites in auditory coincidence detection. Nature 393:268–272. - PubMed
-
- Aizawa H, Hu SC, Bobb K, Balakrishnan K, Ince G, Gurevich I, Cowan M, Ghosh A (2004). Dendrite development regulated by CREST, a calcium-regulated transcriptional activator. Science 303:197–202. - PubMed
-
- Akum BF, Chen M, Gunderson SI, Riefler GM, Scerri-Hansen MM, Firestein BL (2004). Cypin regulates dendrite patterning in hippocampal neurons by promoting microtubule assembly. Nat Neurosci 7:145–152. - PubMed
-
- Ashford JW, Soultanian NS, Zhang SX, Geddes JW (1998). Neuropil threads are collinear with MAP2 immunostaining in neuronal dendrites of Alzheimer brain. J Neuropathol Exp Neurol 57:972–978. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources