

Temporal dynamics and latency patterns of receptor neuron input to the olfactory bulb
- PMID: 16436612
- PMCID: PMC6674558
- DOI: 10.1523/JNEUROSCI.3100-05.2006
Temporal dynamics and latency patterns of receptor neuron input to the olfactory bulb
Abstract
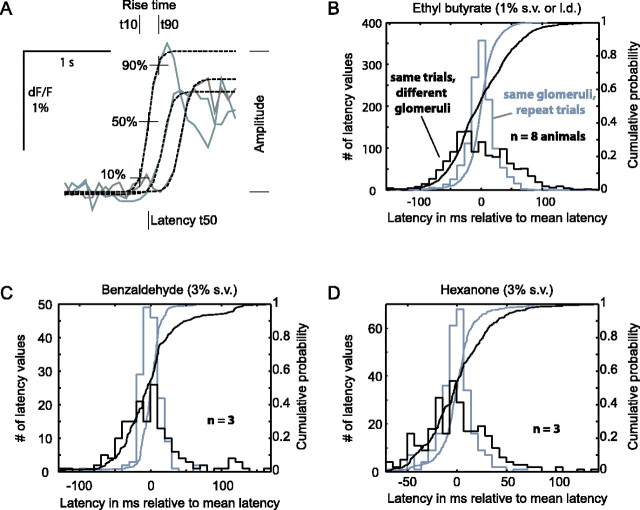
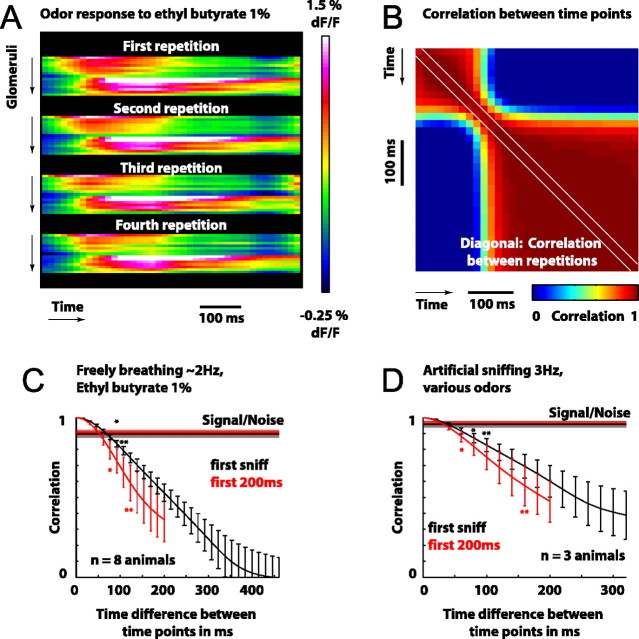
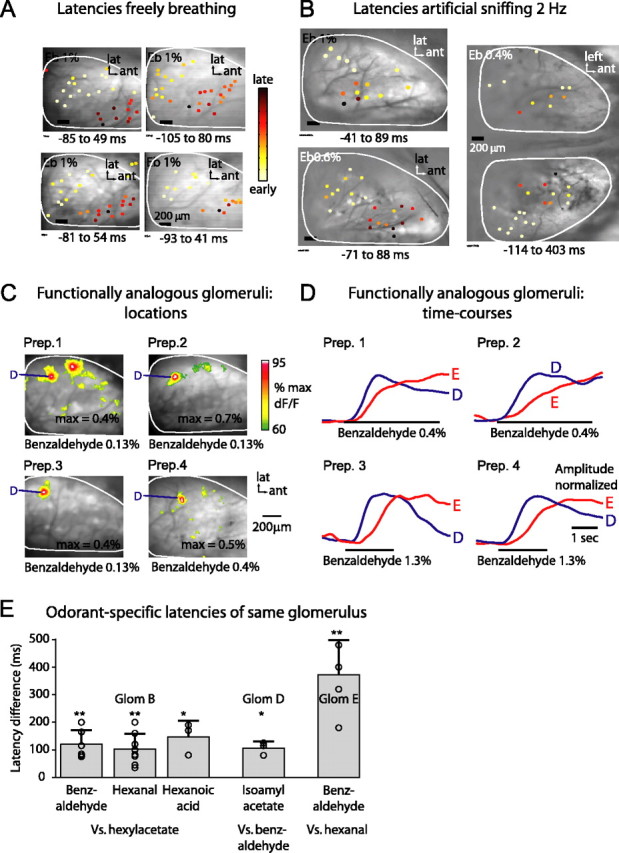
Odorants are first represented in the brain by distributed patterns of activity in the olfactory bulb (OB). Although neurons downstream of sensory inputs respond to odorants with temporally structured activity, sensory inputs to glomeruli are typically described as static maps. Here, we imaged the temporal dynamics of receptor neuron input to the OB with a calcium-sensitive dye in the olfactory receptor nerve terminals in anesthetized mice. We found that diverse, glomerulus- and odorant-dependent temporal dynamics are present even at this initial input stage. Instantaneous spatial patterns of receptor input to glomeruli changed both within and between respiration cycles. Glomerular odorant responses differed in amplitude, latency, rise time, and degree of modulation by sniffing in an odorant-specific manner. Pattern dynamics within the first respiration cycle recurred in a similar manner during consecutive cycles. When sniff rate was increased artificially, pattern dynamics were preserved in the first sniff but were attenuated during subsequent sniffs. Temporal response properties were consistent across individuals on a coarse regional scale and on a fine scale of individual glomeruli. Latency and magnitude of glomerular inputs were only weakly correlated and might therefore convey independent odorant information. These data demonstrate that glomerular maps of primary sensory input to the OB are temporally dynamic. These dynamics may contribute to the representation of odorant information and affect information processing in the central olfactory system of rodents.
Figures




Similar articles
-
Temporal structure of receptor neuron input to the olfactory bulb imaged in behaving rats.J Neurophysiol. 2009 Feb;101(2):1073-88. doi: 10.1152/jn.90902.2008. Epub 2008 Dec 17. J Neurophysiol. 2009. PMID: 19091924 Free PMC article.
-
Representation of odorants by receptor neuron input to the mouse olfactory bulb.Neuron. 2001 Nov 20;32(4):723-35. doi: 10.1016/s0896-6273(01)00506-2. Neuron. 2001. PMID: 11719211
-
Distributed and concentration-invariant spatial representations of odorants by receptor neuron input to the turtle olfactory bulb.J Neurophysiol. 2002 Feb;87(2):1035-45. doi: 10.1152/jn.00522.2001. J Neurophysiol. 2002. PMID: 11826067
-
Olfactory maps and odor images.Curr Opin Neurobiol. 2002 Aug;12(4):387-92. doi: 10.1016/s0959-4388(02)00348-3. Curr Opin Neurobiol. 2002. PMID: 12139985 Review.
-
The olfactory bulb: coding and processing of odor molecule information.Science. 1999 Oct 22;286(5440):711-5. doi: 10.1126/science.286.5440.711. Science. 1999. PMID: 10531048 Review.
Cited by
-
Optical dissection of odor information processing in vivo using GCaMPs expressed in specified cell types of the olfactory bulb.J Neurosci. 2013 Mar 20;33(12):5285-300. doi: 10.1523/JNEUROSCI.4824-12.2013. J Neurosci. 2013. PMID: 23516293 Free PMC article.
-
Functional imaging of cortical feedback projections to the olfactory bulb.Front Neural Circuits. 2014 Jul 3;8:73. doi: 10.3389/fncir.2014.00073. eCollection 2014. Front Neural Circuits. 2014. PMID: 25071454 Free PMC article.
-
Sequence-Based Prediction of Olfactory Receptor Responses.Chem Senses. 2019 Oct 26;44(9):693-703. doi: 10.1093/chemse/bjz059. Chem Senses. 2019. PMID: 31665762 Free PMC article.
-
Diverse populations of intrinsic cholinergic interneurons in the mouse olfactory bulb.Neuroscience. 2012 Jun 28;213:161-78. doi: 10.1016/j.neuroscience.2012.04.024. Epub 2012 Apr 21. Neuroscience. 2012. PMID: 22525133 Free PMC article.
-
The speed of smell: odor-object segregation within milliseconds.PLoS One. 2012;7(4):e36096. doi: 10.1371/journal.pone.0036096. Epub 2012 Apr 27. PLoS One. 2012. PMID: 22558344 Free PMC article.
References
-
- Abraham NM, Spors H, Carleton A, Margrie TW, Kuner T, Schaefer AT (2004). Maintaining accuracy at the expense of speed; stimulus similarity defines odor discrimination time in mice. Neuron 44:865–876. - PubMed
-
- Adrian ED (1950). The electrical activity of the mammalian olfactory bulb. Electroencephalogr Clin Neurophysiol 2:377–388. - PubMed
-
- Ambros-Ingerson J, Granger R, Lynch G (1990). Simulation of paleocortex performs hierarchical clustering. Science 247:1344–1348. - PubMed
-
- Aroniadou-Anderjaska V, Zhou FM, Priest CA, Ennis M, Shipley MT (2000). Tonic and synaptically evoked presynaptic inhibition of sensory input to the rat olfactory bulb via GABAB heteroreceptors. J Neurophysiol 84:1194–1203. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous