

A critical role of erythropoietin receptor in neurogenesis and post-stroke recovery
- PMID: 16436614
- PMCID: PMC6674578
- DOI: 10.1523/JNEUROSCI.4480-05.2006
A critical role of erythropoietin receptor in neurogenesis and post-stroke recovery
Abstract
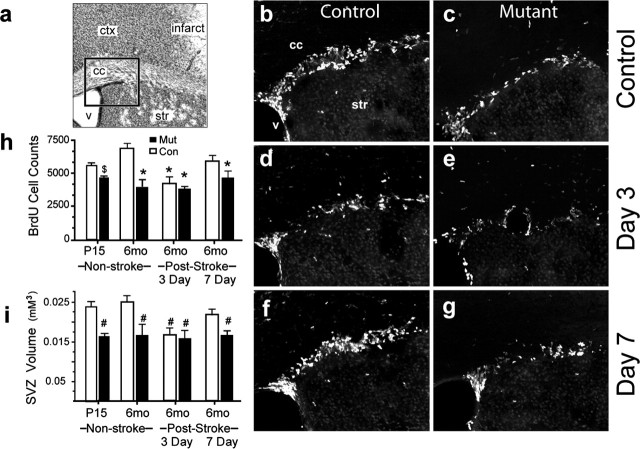
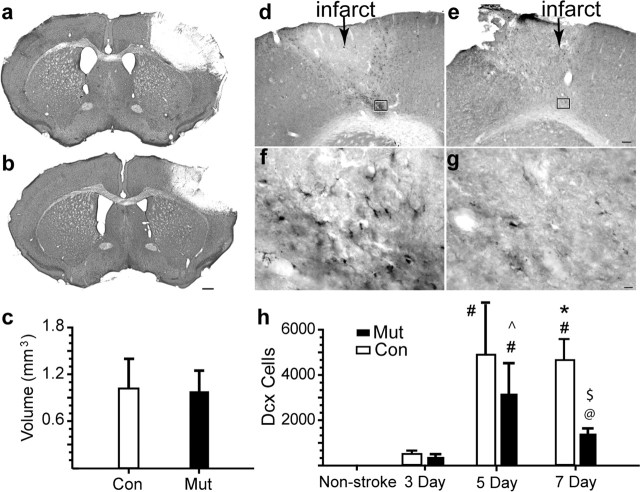
Erythropoietin (EPO) is the principal growth factor regulating the production of red blood cells. Recent studies demonstrated that exogenous EPO acts as a neuroprotectant and regulates neurogenesis. Using a genetic approach, we evaluate the roles of endogenous EPO and its classical receptor (EPOR) in mammalian neurogenesis. We demonstrate severe and identical embryonic neurogenesis defects in animals null for either the Epo or EpoR gene, suggesting that the classical EPOR is essential for EPO action during embryonic neurogenesis. Furthermore, by generating conditional EpoR knock-down animals, we demonstrate that brain-specific deletion of EpoR leads to significantly reduced cell proliferation in the subventricular zone and impaired post-stroke neurogenesis. EpoR conditional knockdown leads to a specific deficit in post-stroke neurogenesis through impaired migration of neuroblasts to the peri-infarct cortex. Our results suggest that both EPO and EPOR are essential for early embryonic neural development and that the classical EPOR is important for adult neurogenesis and for migration of regenerating neurons during post-injury recovery.
Figures


References
-
- Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O (2002). Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med 8:963–970. - PubMed
-
- Bernaudin M, Marti HH, Roussel S, Divoux D, Nouvelot A, MacKenzie ET, Petit E (1999). A potential role for erythropoietin in focal permanent cerebral ischemia in mice. J Cereb Blood Flow Metab 19:643–651. - PubMed
-
- Brines M, Grasso G, Fiordaliso F, Sfacteria A, Ghezzi P, Fratelli M, Latini R, Xie QW, Smart J, Su-Rick CJ, Pobre E, Diaz D, Gomez D, Hand C, Coleman T, Cerami A (2004). Erythropoietin mediates tissue protection through an erythropoietin and common beta-subunit heteroreceptor. Proc Natl Acad Sci USA 101:14907–14912. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials