

Granzyme A, which causes single-stranded DNA damage, targets the double-strand break repair protein Ku70
- PMID: 16440001
- PMCID: PMC1456912
- DOI: 10.1038/sj.embor.7400622
Granzyme A, which causes single-stranded DNA damage, targets the double-strand break repair protein Ku70
Abstract
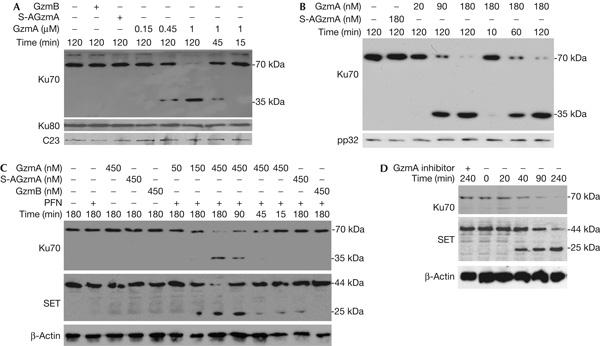
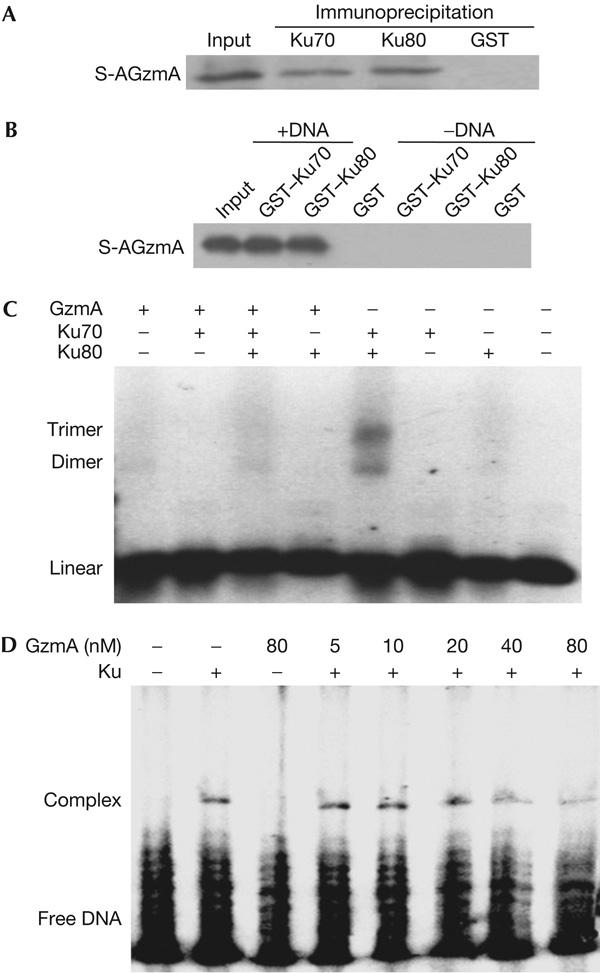
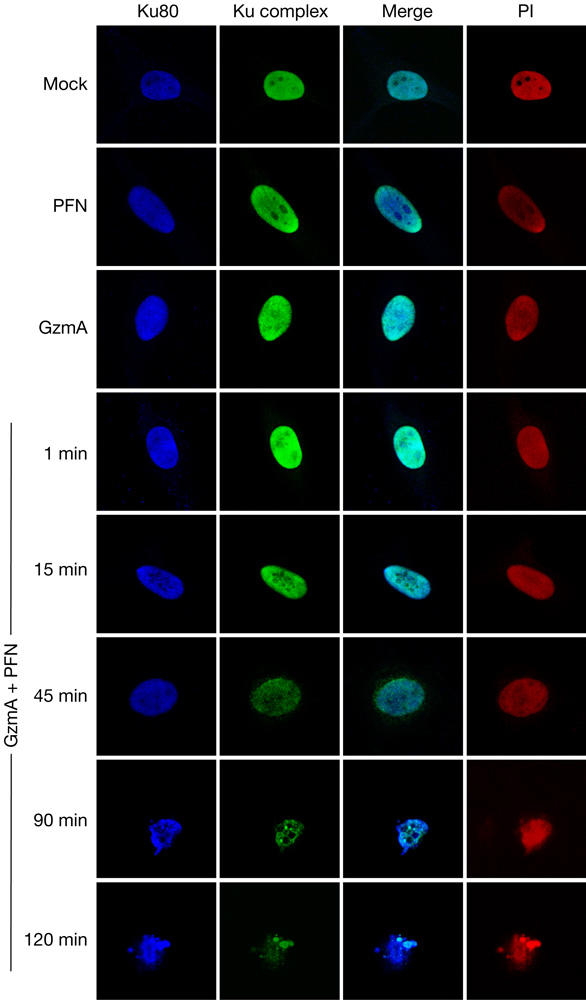
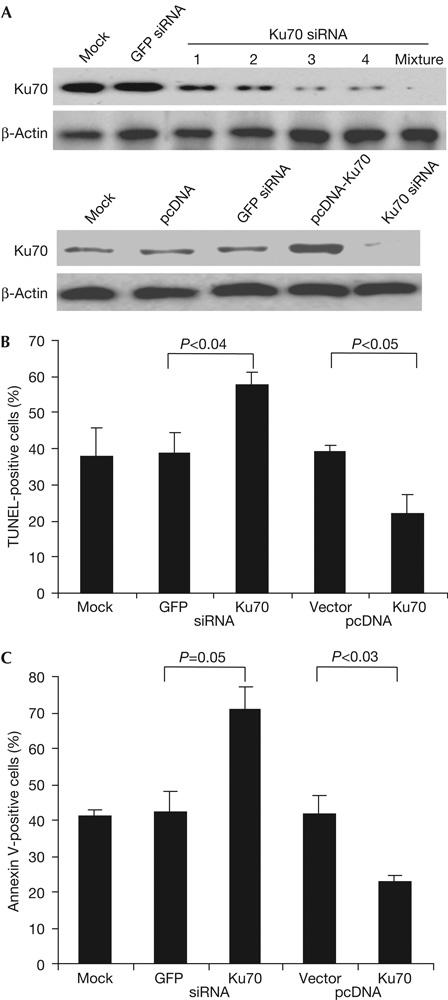
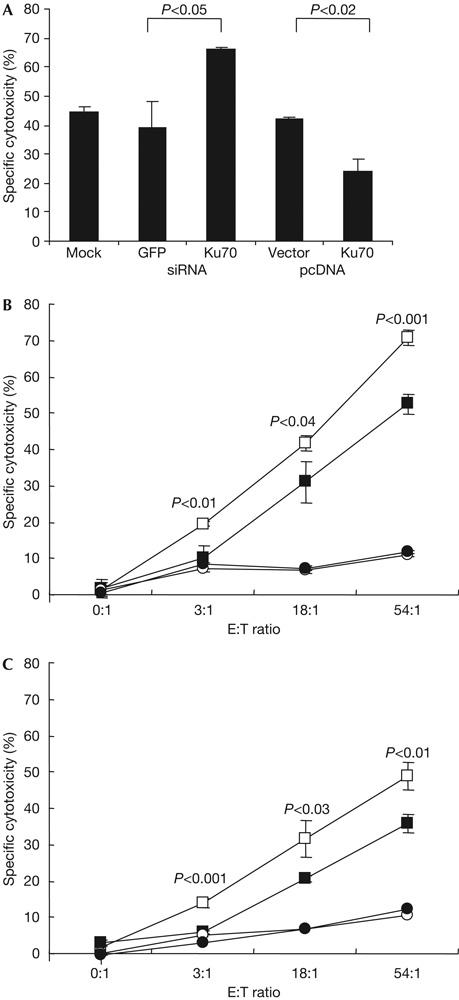
Granzyme A (GzmA) induces caspase-independent cell death with morphological features of apoptosis. Here, we show that GzmA at nanomolar concentrations cleaves Ku70, a key double-strand break repair (DSBR) protein, in target cells. Ku70 is cut after Arg(301), disrupting Ku complex binding to DNA. Cleaving Ku70 facilitates GzmA-mediated cell death, as silencing Ku70 by RNA interference increases DNA damage and cell death by GzmB cluster-deficient cytotoxic T lymphocytes or by GzmA and perforin, whereas Ku70 overexpression has the opposite effect. Ku70 has two known antiapoptotic effects-facilitating DSBR and sequestering bax to prevent its translocation to mitochondria. However, GzmA triggers single-stranded, not double-stranded, DNA damage, and GzmA-induced cell death does not involve bax. Therefore, Ku70 has other antiapoptotic functions in GzmA-induced cell death, which are blocked when GzmA proteolyses Ku70.
Figures
Similar articles
-
Granzyme A activates another way to die.Immunol Rev. 2010 May;235(1):93-104. doi: 10.1111/j.0105-2896.2010.00902.x. Immunol Rev. 2010. PMID: 20536557 Free PMC article. Review.
-
Cleaving the oxidative repair protein Ape1 enhances cell death mediated by granzyme A.Nat Immunol. 2003 Feb;4(2):145-53. doi: 10.1038/ni885. Epub 2003 Jan 13. Nat Immunol. 2003. PMID: 12524539
-
Granzyme A activates an endoplasmic reticulum-associated caspase-independent nuclease to induce single-stranded DNA nicks.J Biol Chem. 2001 Nov 16;276(46):43285-93. doi: 10.1074/jbc.M108137200. Epub 2001 Sep 12. J Biol Chem. 2001. PMID: 11555662
-
In vitro- and ex vivo-derived cytolytic leukocytes from granzyme A x B double knockout mice are defective in granule-mediated apoptosis but not lysis of target cells.J Exp Med. 1997 Nov 17;186(10):1781-6. doi: 10.1084/jem.186.10.1781. J Exp Med. 1997. PMID: 9362539 Free PMC article.
-
Nuclear war: the granzyme A-bomb.Curr Opin Immunol. 2003 Oct;15(5):553-9. doi: 10.1016/s0952-7915(03)00108-0. Curr Opin Immunol. 2003. PMID: 14499264 Review.
Cited by
-
Modulation of Inflammation by Extracellular Granzyme A.Front Immunol. 2020 May 19;11:931. doi: 10.3389/fimmu.2020.00931. eCollection 2020. Front Immunol. 2020. PMID: 32508827 Free PMC article. Review.
-
PARP-1 cleavage fragments: signatures of cell-death proteases in neurodegeneration.Cell Commun Signal. 2010 Dec 22;8:31. doi: 10.1186/1478-811X-8-31. Cell Commun Signal. 2010. PMID: 21176168 Free PMC article.
-
Accumulation of CD28null Senescent T-Cells Is Associated with Poorer Outcomes in COVID19 Patients.Biomolecules. 2021 Sep 29;11(10):1425. doi: 10.3390/biom11101425. Biomolecules. 2021. PMID: 34680058 Free PMC article. Review.
-
Granzyme A activates another way to die.Immunol Rev. 2010 May;235(1):93-104. doi: 10.1111/j.0105-2896.2010.00902.x. Immunol Rev. 2010. PMID: 20536557 Free PMC article. Review.
-
Ku80-deleted cells are defective at base excision repair.Mutat Res. 2013 May-Jun;745-746:16-25. doi: 10.1016/j.mrfmmm.2013.03.010. Epub 2013 Apr 6. Mutat Res. 2013. PMID: 23567907 Free PMC article.
References
-
- Andrade F, Roy S, Nicholson D, Thornberry N, Rosen A, Casciola-Rosen L (1998) Granzyme B directly and efficiently cleaves several downstream caspase substrates: implications for CTL-induced apoptosis. Immunity 8: 451–460 - PubMed
-
- Bell JK, Goetz DH, Mahrus S, Harris JL, Fletterick RJ, Craik CS (2003) The oligomeric structure of human granzyme A is a determinant of its extended substrate specificity. Nat Struct Biol 10: 527–534 - PubMed
-
- Beresford PJ, Xia Z, Greenberg AH, Lieberman J (1999) Granzyme A loading induces rapid cytolysis and a novel form of DNA damage independently of caspase activation. Immunity 10: 585–594 - PubMed
-
- Beresford PJ, Zhang D, Oh DY, Fan Z, Greer EL, Russo ML, Jaju M, Lieberman J (2001) Granzyme A activates an endoplasmic reticulum-associated caspase-independent nuclease to induce single-stranded DNA nicks. J Biol Chem 276: 43285–43293 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
