

Genetic analysis of the cytoplasmic dynein subunit families
- PMID: 16440056
- PMCID: PMC1331979
- DOI: 10.1371/journal.pgen.0020001
Genetic analysis of the cytoplasmic dynein subunit families
Abstract
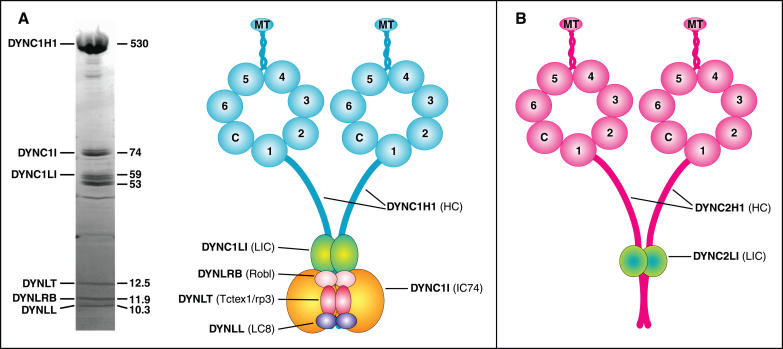
Cytoplasmic dyneins, the principal microtubule minus-end-directed motor proteins of the cell, are involved in many essential cellular processes. The major form of this enzyme is a complex of at least six protein subunits, and in mammals all but one of the subunits are encoded by at least two genes. Here we review current knowledge concerning the subunits, their interactions, and their functional roles as derived from biochemical and genetic analyses. We also carried out extensive database searches to look for new genes and to clarify anomalies in the databases. Our analysis documents evolutionary relationships among the dynein subunits of mammals and other model organisms, and sheds new light on the role of this diverse group of proteins, highlighting the existence of two cytoplasmic dynein complexes with distinct cellular roles.
Figures

Similar articles
-
DYNLT (Tctex-1) forms a tripartite complex with dynein intermediate chain and RagA, hence linking this small GTPase to the dynein motor.FEBS J. 2015 Oct;282(20):3945-58. doi: 10.1111/febs.13388. Epub 2015 Aug 20. FEBS J. 2015. PMID: 26227614
-
Dynein interacts with the neural cell adhesion molecule (NCAM180) to tether dynamic microtubules and maintain synaptic density in cortical neurons.J Biol Chem. 2013 Sep 27;288(39):27812-24. doi: 10.1074/jbc.M113.465088. Epub 2013 Aug 19. J Biol Chem. 2013. PMID: 23960070 Free PMC article.
-
Reconstitution of the human cytoplasmic dynein complex.Proc Natl Acad Sci U S A. 2012 Dec 18;109(51):20895-900. doi: 10.1073/pnas.1210573110. Epub 2012 Dec 4. Proc Natl Acad Sci U S A. 2012. PMID: 23213255 Free PMC article.
-
Molecular organization and force-generating mechanism of dynein.FEBS J. 2011 Sep;278(17):2964-79. doi: 10.1111/j.1742-4658.2011.08253.x. Epub 2011 Aug 8. FEBS J. 2011. PMID: 21777385 Review.
-
Structure and Function of Dynein's Non-Catalytic Subunits.Cells. 2024 Feb 11;13(4):330. doi: 10.3390/cells13040330. Cells. 2024. PMID: 38391943 Free PMC article. Review.
Cited by
-
Dlic1 deficiency impairs ciliogenesis of photoreceptors by destabilizing dynein.Cell Res. 2013 Jun;23(6):835-50. doi: 10.1038/cr.2013.59. Epub 2013 Apr 30. Cell Res. 2013. PMID: 23628724 Free PMC article.
-
Dynein Light Chain 1 (DYNLT1) Interacts with Normal and Oncogenic Nucleoporins.PLoS One. 2013 Jun 26;8(6):e67032. doi: 10.1371/journal.pone.0067032. Print 2013. PLoS One. 2013. PMID: 23840580 Free PMC article.
-
Distinct functional roles of cytoplasmic dynein defined by the intermediate chain isoforms.Exp Cell Res. 2015 May 15;334(1):54-60. doi: 10.1016/j.yexcr.2014.12.013. Epub 2015 Jan 6. Exp Cell Res. 2015. PMID: 25576383 Free PMC article. Review.
-
Mechanical dynamics in live cells and fluorescence-based force/tension sensors.Biochim Biophys Acta. 2015 Aug;1853(8):1889-904. doi: 10.1016/j.bbamcr.2015.05.001. Epub 2015 May 6. Biochim Biophys Acta. 2015. PMID: 25958335 Free PMC article. Review.
-
Few Fixed Variants between Trophic Specialist Pupfish Species Reveal Candidate Cis-Regulatory Alleles Underlying Rapid Craniofacial Divergence.Mol Biol Evol. 2021 Jan 23;38(2):405-423. doi: 10.1093/molbev/msaa218. Mol Biol Evol. 2021. PMID: 32877534 Free PMC article.
References
-
- Gibbons IR. Chemical dissection of cilia. Arch Biol (Liege) 1965;76:317–352. - PubMed
-
- Paschal BM, King SM, Moss AG, Collins CA, Vallee RB, et al. Isolated flagellar outer arm dynein translocates brain microtubules in vitro. Nature. 1987;330:672–674. - PubMed
-
- Paschal BM, Vallee RB. Retrograde transport by the microtubule-associated protein MAP 1C. Nature. 1987;330:181–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
