

Glucocorticoids exacerbate obesity and insulin resistance in neuron-specific proopiomelanocortin-deficient mice
- PMID: 16440060
- PMCID: PMC1350998
- DOI: 10.1172/JCI25243
Glucocorticoids exacerbate obesity and insulin resistance in neuron-specific proopiomelanocortin-deficient mice
Erratum in
- J Clin Invest. 2006 Mar;116(3):842
Abstract
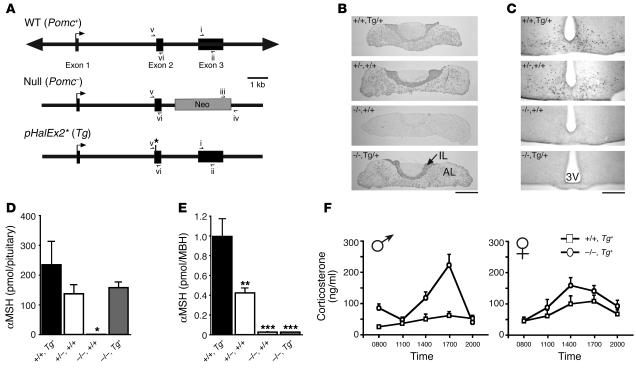
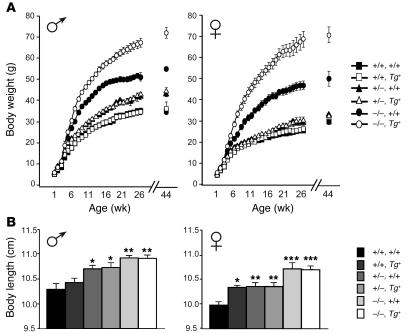
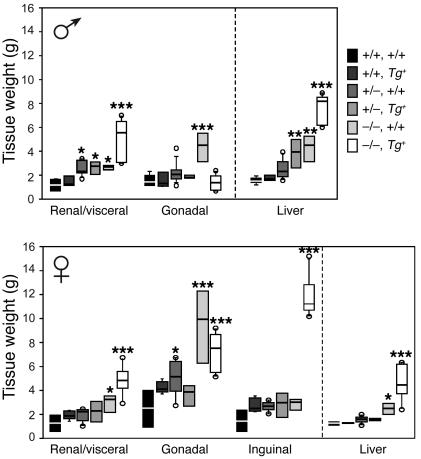
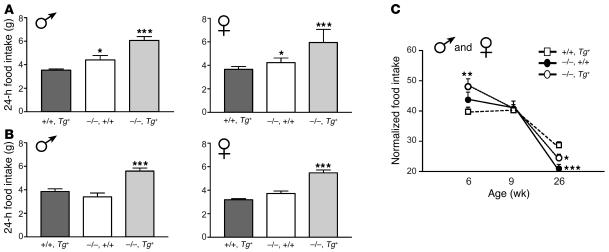
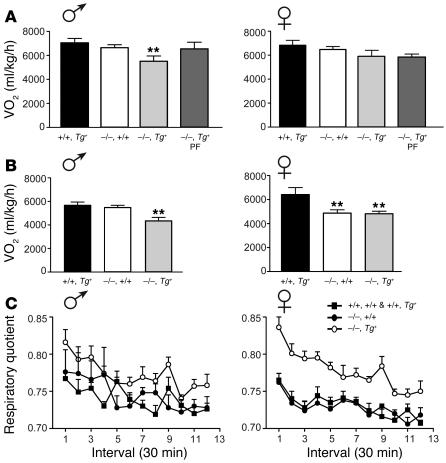
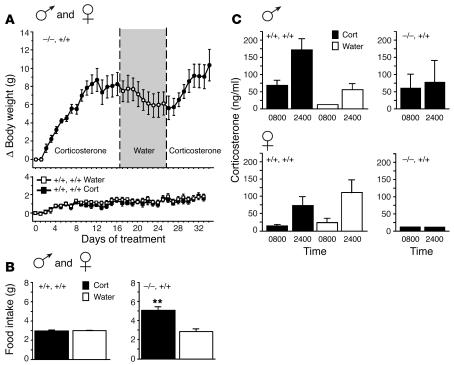
Null mutations of the proopiomelanocortin gene (Pomc) cause obesity in humans and rodents, but the contributions of central versus pituitary POMC deficiency are not fully established. To elucidate these roles, we introduced a POMC transgene (Tg) that selectively restored peripheral melanocortin and corticosterone secretion in Pomc mice. Rather than improving energy balance, the genetic replacement of pituitary POMC in PomcTg mice aggravated their metabolic syndrome with increased caloric intake and feed efficiency, reduced oxygen consumption, increased subcutaneous, visceral, and hepatic fat, and severe insulin resistance. Pair-feeding of PomcTg mice to the daily intake of lean controls normalized their rate of weight gain but did not abolish obesity, indicating that hyperphagia is a major but not sole determinant of the phenotype. Replacement of corticosterone in the drinking water of Pomc mice recapitulated the hyperphagia, excess weight gain and fat accumulation, and hyperleptinemia characteristic of genetically rescued PomcTg mice. These data demonstrate that CNS POMC peptides play a critical role in energy homeostasis that is not substituted by peripheral POMC. Restoration of pituitary POMC expression to create a de facto neuronal POMC deficiency exacerbated the development of obesity, largely via glucocorticoid modulation of appetite, metabolism, and energy partitioning.
Figures

References
-
- Isomaa B. A major health hazard: the metabolic syndrome. Life Sci. 2003;73:2395–2411. - PubMed
-
- Havel PJ. Peripheral signals conveying metabolic information to the brain: short-term and long-term regulation of food intake and energy homeostasis. Exp. Biol. Med. 2001;226:963–977. - PubMed
-
- Dallman MF, et al. A spoonful of sugar: feedback signals of energy stores and corticosterone regulate responses to chronic stress. Physiol. Behav. 2003;79:3–12. - PubMed
-
- Low MJ. Role of proopiomelanocortin neurons and peptides in the regulation of energy homeostasis. J. Endocrinol. Invest. 2004;27:95–100. - PubMed
-
- Ellacott KL, Cone RD. The central melanocortin system and the integration of short- and long-term regulators of energy homeostasis. Recent Prog. Horm. Res. 2004;59:395–408. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
