

A method for determining the dependence of calcium oscillations on inositol trisphosphate oscillations
- PMID: 16446452
- PMCID: PMC1413622
- DOI: 10.1073/pnas.0506135103
A method for determining the dependence of calcium oscillations on inositol trisphosphate oscillations
Abstract
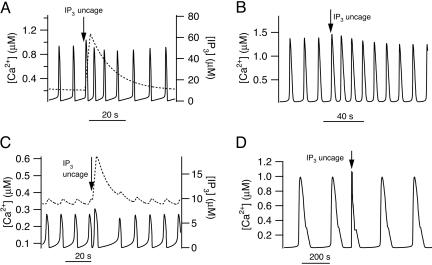
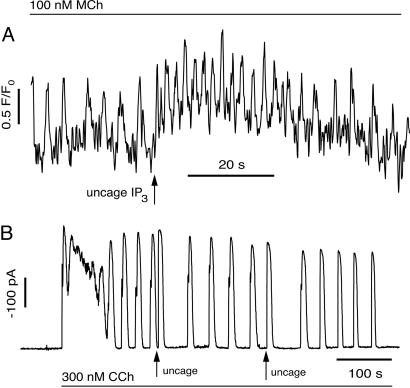
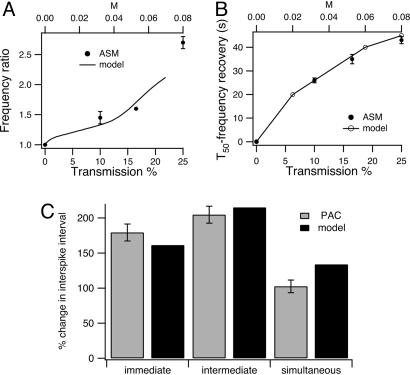
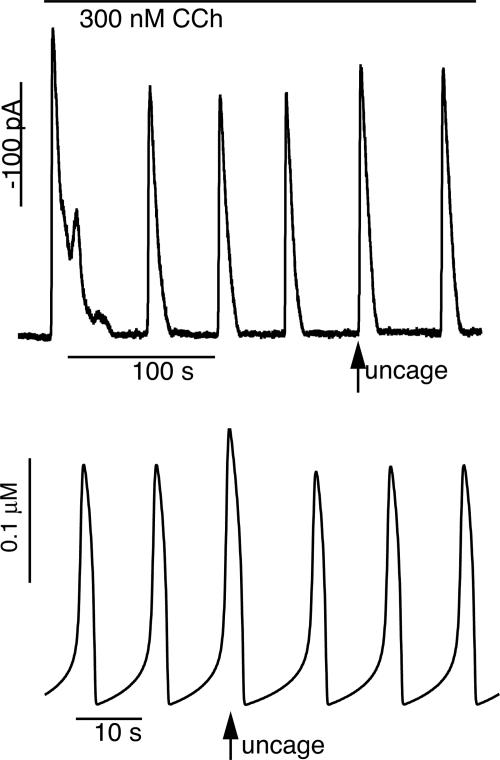
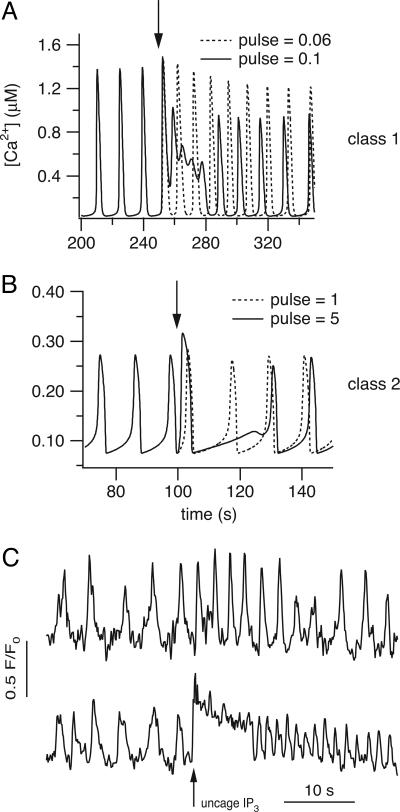
In some cell types, oscillations in the concentration of free intracellular calcium ([Ca2+]) are accompanied by oscillations in the concentration of inositol 1,4,5-trisphosphate ([IP3]). However, in most cell types it is still an open question as to whether oscillations in [IP3] are necessary for Ca2+ oscillations in vivo, or whether they merely follow passively. Using a wide range of models, we show that the response to an artificially applied pulse of IP3 can be used to distinguish between these two cases. Hence, we show that muscarinic receptor-mediated, long-period Ca2+ oscillations in pancreatic acinar cells depend on [IP3] oscillations, whereas short-period Ca2+ oscillations in airway smooth muscle do not.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
