

HEN1 recognizes 21-24 nt small RNA duplexes and deposits a methyl group onto the 2' OH of the 3' terminal nucleotide
- PMID: 16449203
- PMCID: PMC1356533
- DOI: 10.1093/nar/gkj474
HEN1 recognizes 21-24 nt small RNA duplexes and deposits a methyl group onto the 2' OH of the 3' terminal nucleotide
Abstract
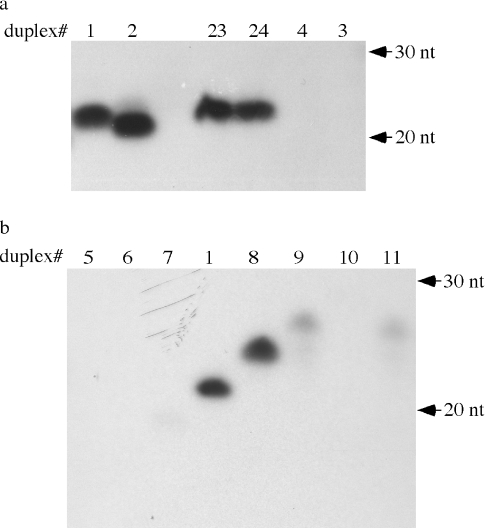
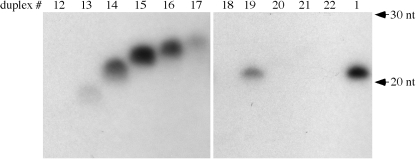
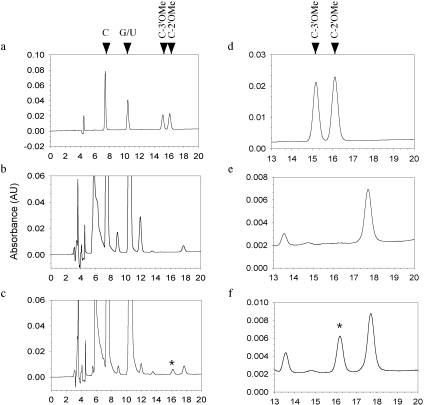
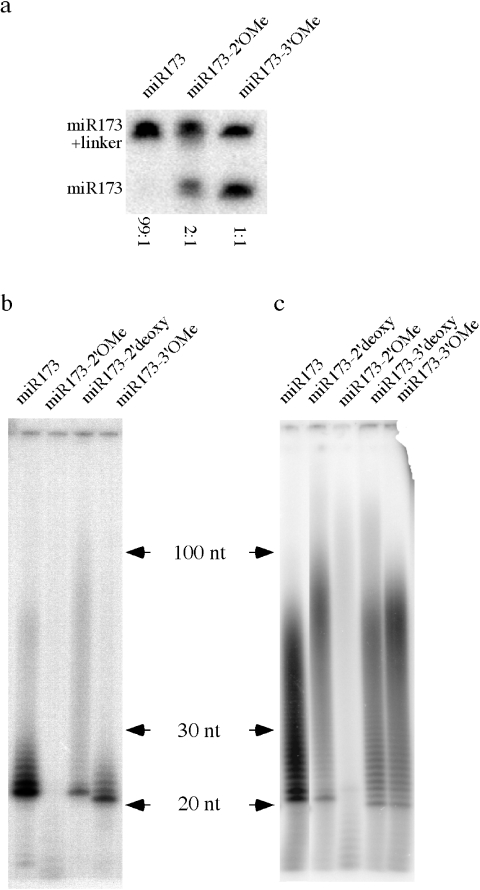
microRNAs (miRNAs) and small interfering RNAs (siRNAs) in plants bear a methyl group on the ribose of the 3' terminal nucleotide. We showed previously that the methylation of miRNAs and siRNAs requires the protein HEN1 in vivo and that purified HEN1 protein methylates miRNA/miRNA* duplexes in vitro. In this study, we show that HEN1 methylates both miRNA/miRNA* and siRNA/siRNA* duplexes in vitro with a preference for 21-24 nt RNA duplexes with 2 nt overhangs. We also demonstrate that HEN1 deposits the methyl group on to the 2' OH of the 3' terminal nucleotide. Among various modifications that can occur on the ribose of the terminal nucleotide, such as 2'-deoxy, 3'-deoxy, 2'-O-methyl and 3'-O-methyl, only 2'-O-methyl on a small RNA inhibits the activity of yeast poly(A) polymerase (PAP). These findings indicate that HEN1 specifically methylates miRNAs and siRNAs and implicate the importance of the 2'-O-methyl group in the biology of RNA silencing.
Figures
References
-
- Hammond S.M. Dicing and slicing: the core machinery of the RNA interference pathway. FEBS Lett. 2005;579:5822–5829. - PubMed
-
- Matzke M.A., Birchler J.A. RNAi-mediated pathways in the nucleus. Nature Rev. Genet. 2005;6:24–35. - PubMed
-
- Morel J.B., Mourrain P., Beclin C., Vaucheret H. DNA methylation and chromatin structure affect transcriptional and post-transcriptional transgene silencing in Arabidopsis. Curr. Biol. 2000;10:1591–1594. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
