

Antigen receptor-mediated changes in glucose metabolism in B lymphocytes: role of phosphatidylinositol 3-kinase signaling in the glycolytic control of growth
- PMID: 16449529
- PMCID: PMC1895797
- DOI: 10.1182/blood-2005-12-4788
Antigen receptor-mediated changes in glucose metabolism in B lymphocytes: role of phosphatidylinositol 3-kinase signaling in the glycolytic control of growth
Abstract
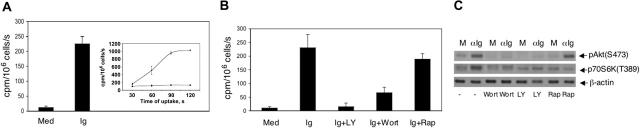
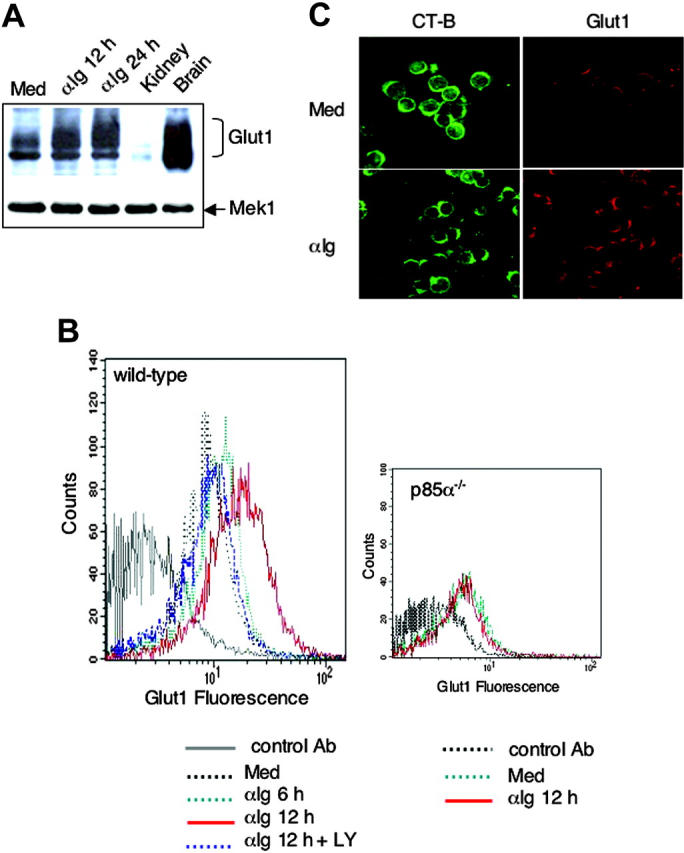
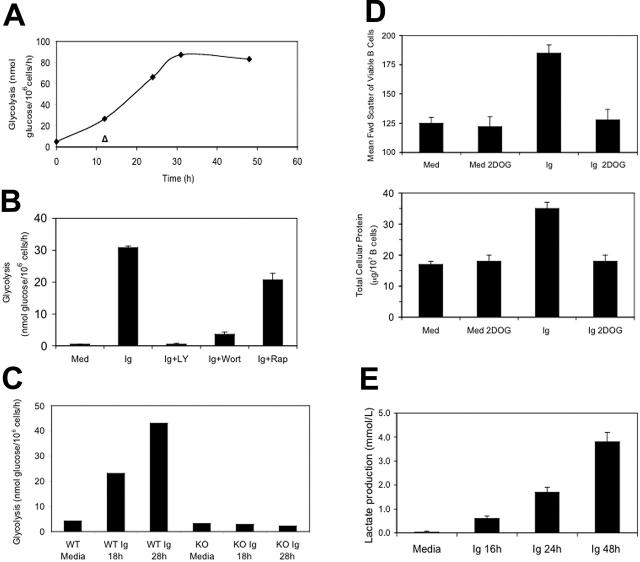
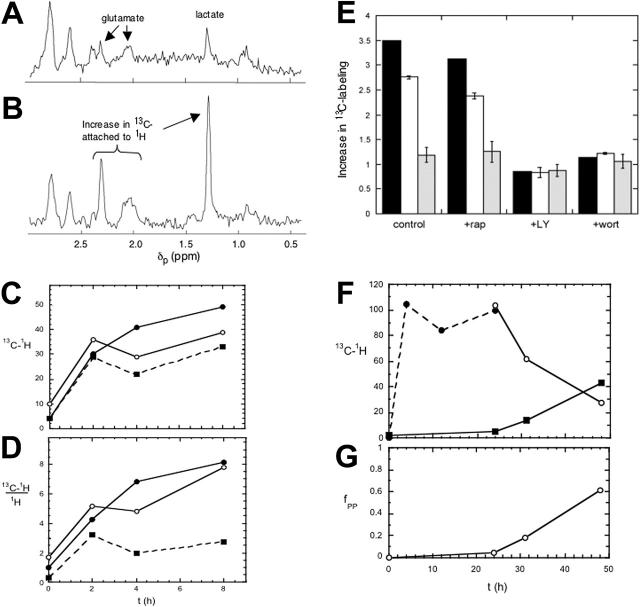
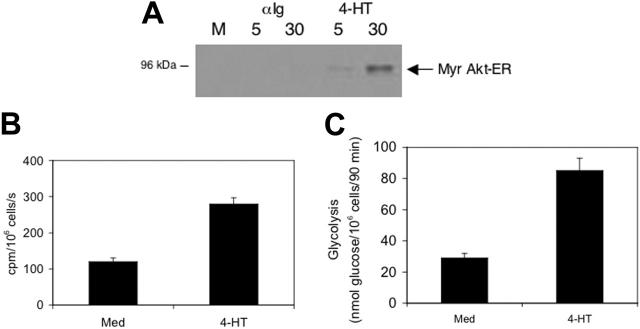
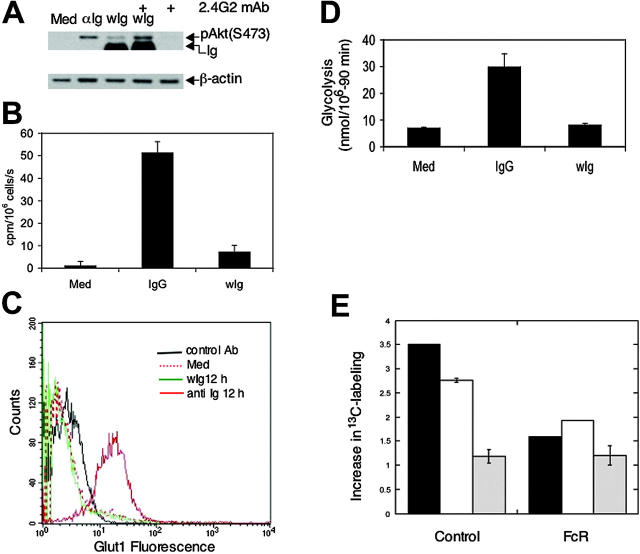
The bioenergetic response of B lymphocytes is subject to rapid changes following antigen encounter in order to provide ATP and anabolic precursors necessary to support growth. However, the pathways involved in glucose acquisition and metabolism are unknown. We find that B lymphocytes rapidly increase glucose uptake and glycolysis following B-cell antigen receptor (BCR) crosslinking. Inhibition of glycolysis blocks BCR-mediated growth. Prior to S-phase entry, glucose metabolism shifts from primarily glycolytic to include the pentose phosphate pathway. BCR-induced glucose utilization is dependent upon phosphatidylinositol 3-kinase (PI-3K) activity as evidenced by inhibition of glucose uptake and glycolysis with LY294002 treatment of normal B cells and impaired glucose utilization in B cells deficient in the PI-3K regulatory subunit p85alpha. Activation of Akt is sufficient to increase glucose utilization in B cells. We find that glucose utilization is inhibited by coengagement of the BCR and FcgammaRIIB, suggesting that limiting glucose metabolism may represent an important mechanism underlying FcgammaRIIB-mediated growth arrest. Taken together, these findings demonstrate that both growth-promoting BCR signaling and growth-inhibitory FcgammaRIIB signaling modulate glucose energy metabolism. Manipulation of these pathways may prove to be useful in the treatment of lymphoproliferative disorders, wherein clonal expansion of B lymphocytes plays a role.
Figures
References
-
- DeFranco AL, Raveche ES, Paul WE. Separate control of B lymphocyte early activation and proliferation in response to anti-IgM antibodies. J Immunol. 1985;135: 87-94. - PubMed
-
- Sieckmann DG. The use of anti-immunoglobulins to induce a signal for cell division in B lymphocytes via their membrane IgM and IgD. Immunol Rev. 1980;52: 181-210. - PubMed
-
- Schuhmacher M, Staege MS, Pajic A, et al. Control of cell growth by c-Myc in the absence of cell division. Curr Biol. 1999;9: 1255-1258. - PubMed
-
- Buttgereit F, Burmester GR, Brand MD. Bioenergetics of immune functions: fundamental and therapeutic aspects. Immunol Today. 2000;21: 192-199. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
