

Comprehensive mutational analysis of yeast DEXD/H box RNA helicases involved in large ribosomal subunit biogenesis
- PMID: 16449635
- PMCID: PMC1367183
- DOI: 10.1128/MCB.26.4.1195-1208.2006
Comprehensive mutational analysis of yeast DEXD/H box RNA helicases involved in large ribosomal subunit biogenesis
Abstract
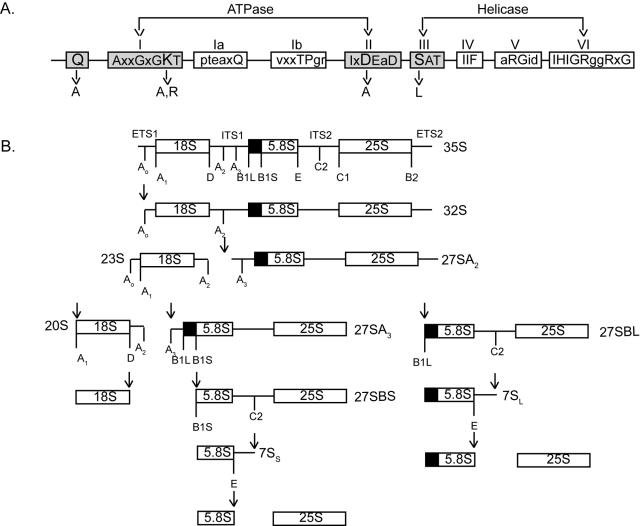
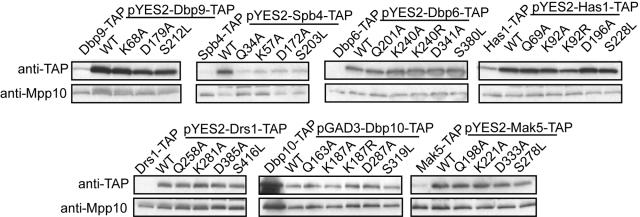
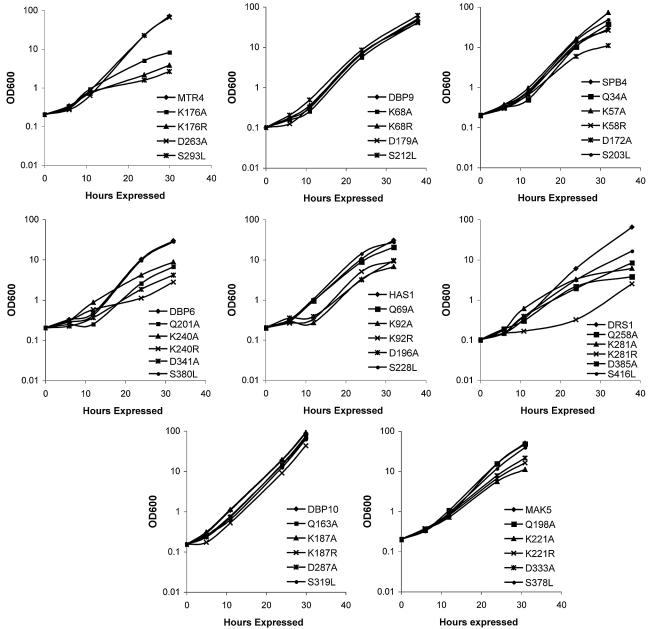
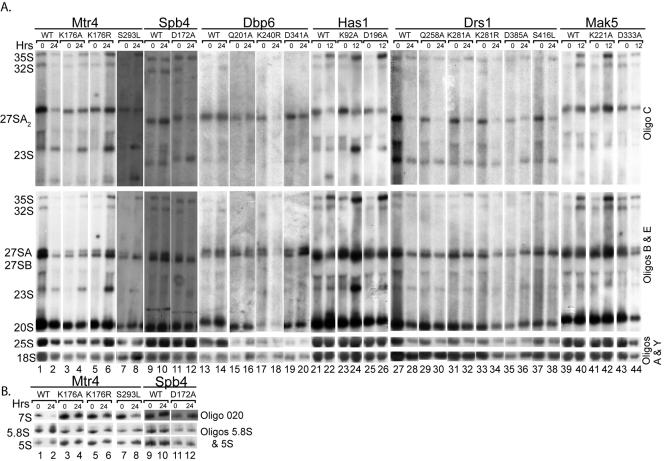
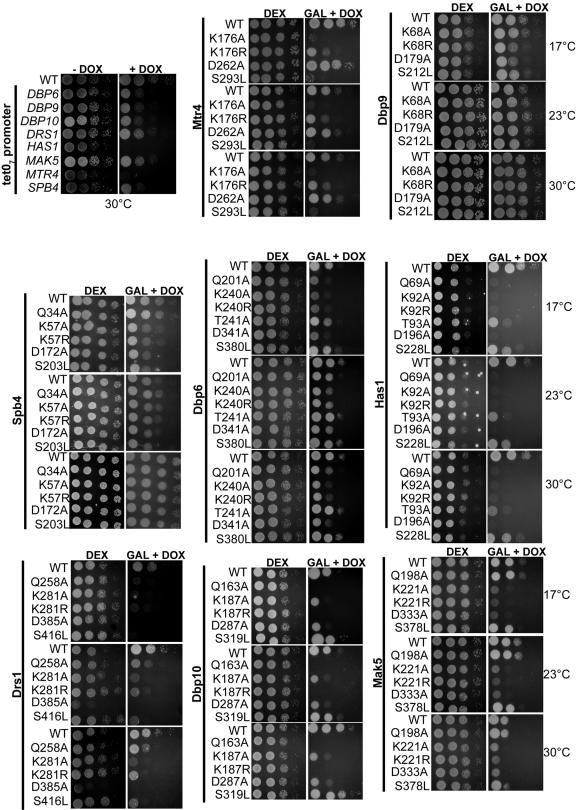
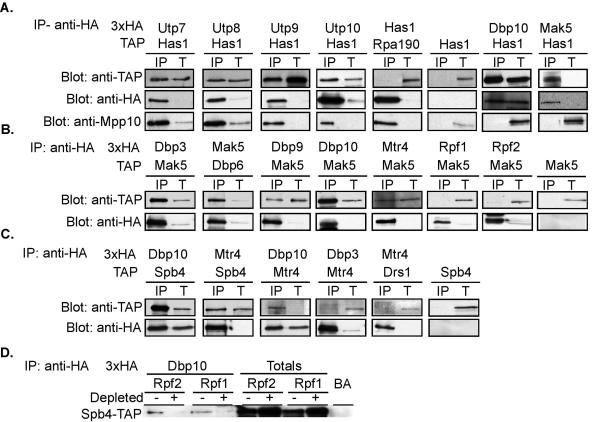
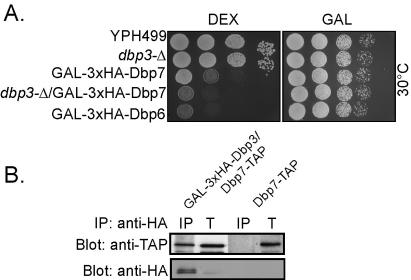
DEXD/H box putative RNA helicases are required for pre-rRNA processing in Saccharomyces cerevisiae, although their exact roles and substrates are unknown. To characterize the significance of the conserved motifs for helicase function, a series of five mutations were created in each of the eight essential RNA helicases (Has1, Dbp6, Dbp10, Mak5, Mtr4, Drs1, Spb4, and Dbp9) involved in 60S ribosomal subunit biogenesis. Each mutant helicase was screened for the ability to confer dominant negative growth defects and for functional complementation. Different mutations showed different degrees of growth inhibition among the helicases, suggesting that the conserved regions do not function identically in vivo. Mutations in motif I and motif II (the DEXD/H box) often conferred dominant negative growth defects, indicating that these mutations do not interfere with substrate binding. In addition, mutations in the putative unwinding domains (motif III) demonstrated that conserved amino acids are often not essential for function. Northern analysis of steady-state RNA from strains expressing mutant helicases showed that the dominant negative mutations also altered pre-rRNA processing. Coimmunoprecipitation experiments indicated that some RNA helicases associated with each other. In addition, we found that yeasts disrupted in expression of the two nonessential RNA helicases, Dbp3 and Dbp7, grew worse than when either one alone was disrupted.
Figures

References
-
- Belli, G., E. Gari, M. Aldea, and E. Herrero. 1998. Functional analysis of yeast essential genes using a promoter-substitution cassette and the tetracycline-regulatable dual expression system. Yeast 14:1127-1138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases