

Chromatin decondensation and nuclear reprogramming by nucleoplasmin
- PMID: 16449640
- PMCID: PMC1367201
- DOI: 10.1128/MCB.26.4.1259-1271.2006
Chromatin decondensation and nuclear reprogramming by nucleoplasmin
Erratum in
- Mol Cell Biol. 2007 Sep;27(18):6580
Abstract
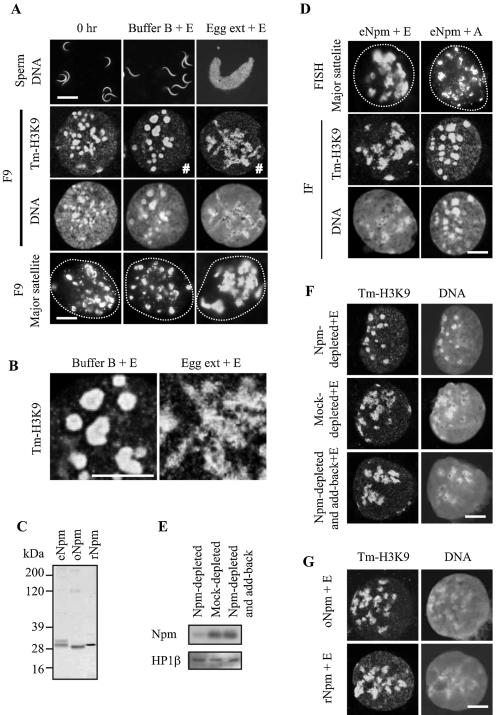
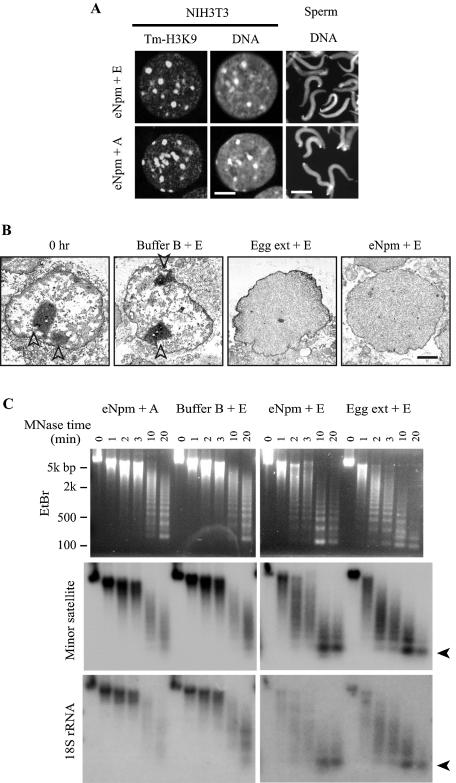
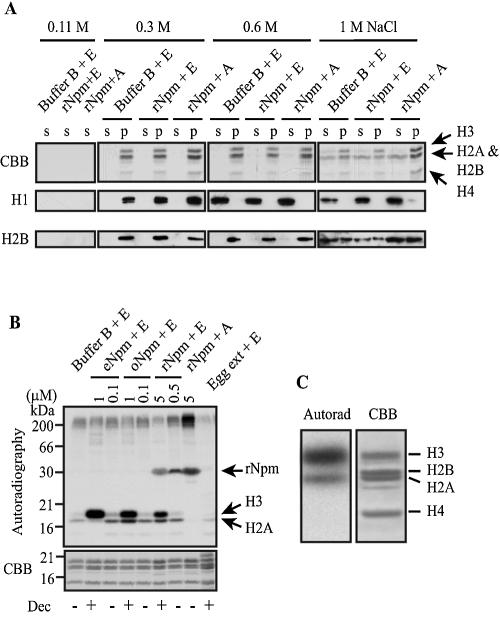
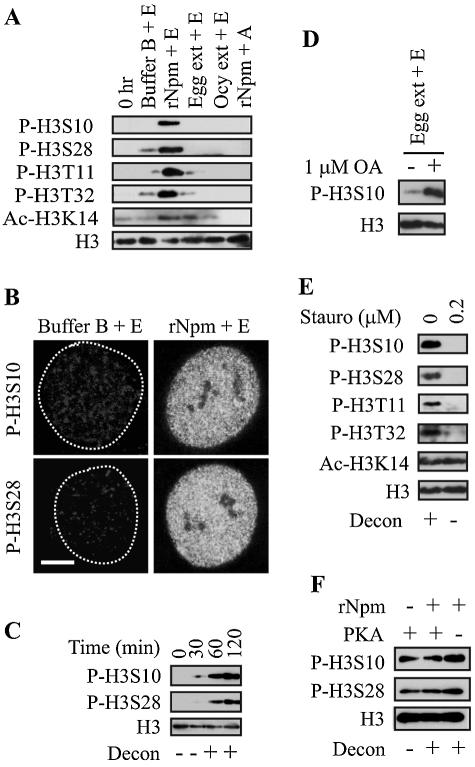
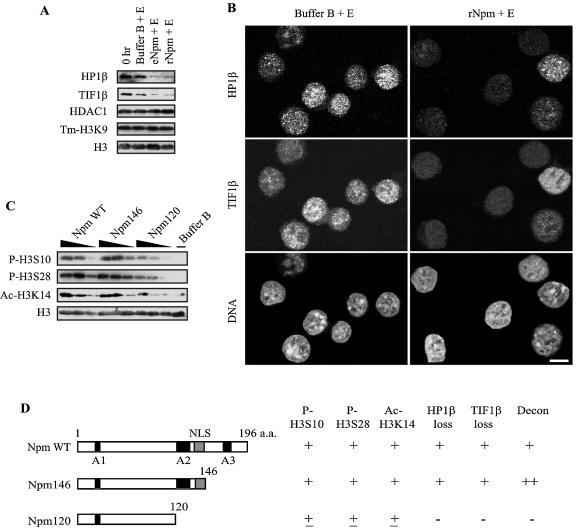
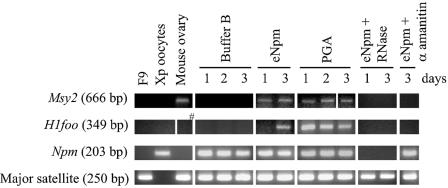
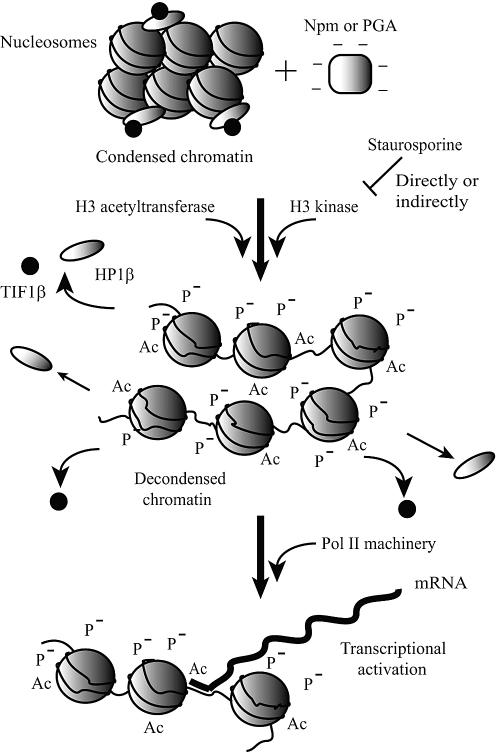
Somatic cell nuclear cloning has repeatedly demonstrated striking reversibility of epigenetic regulation of cell differentiation. Upon injection into eggs, the donor nuclei exhibit global chromatin decondensation, which might contribute to reprogramming the nuclei by derepressing dormant genes. Decondensation of sperm chromatin in eggs is explained by the replacement of sperm-specific histone variants with egg-type histones by the egg protein nucleoplasmin (Npm). However, little is known about the mechanisms of chromatin decondensation in somatic nuclei that do not contain condensation-specific histone variants. Here we found that Npm could widely decondense chromatin in undifferentiated mouse cells without overt histone exchanges but with specific epigenetic modifications that are relevant to open chromatin structure. These modifications included nucleus-wide multiple histone H3 phosphorylation, acetylation of Lys 14 in histone H3, and release of heterochromatin proteins HP1beta and TIF1beta from the nuclei. The protein kinase inhibitor staurosporine inhibited chromatin decondensation and these epigenetic modifications with the exception of H3 acetylation, potentially linking these chromatin events. At the functional level, Npm pretreatment of mouse nuclei facilitated activation of four oocyte-specific genes from the nuclei injected into Xenopus laevis oocytes. Future molecular elucidation of chromatin decondensation by Npm will significantly contribute to our understanding of the plasticity of cell differentiation.
Figures
References
-
- Ajiro, K., K. Yoda, K. Utsumi, and Y. Nishikawa. 1996. Alteration of cell cycle-dependent histone phosphorylations by okadaic acid. Induction of mitosis-specific H3 phosphorylation and chromatin condensation in mammalian interphase cells. J. Biol. Chem. 271:13197-13201. - PubMed
-
- Arnan, C., N. Saperas, C. Prieto, M. Chiva, and J. Ausio. 2003. Interaction of nucleoplasmin with core histones. J. Biol. Chem. 278:31319-31324. - PubMed
-
- Bannister, A. J., P. Zegerman, J. F. Partridge, E. A. Miska, J. O. Thomas, R. C. Allshire, and T. Kouzarides. 2001. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410:120-124. - PubMed
-
- Banuelos, S., A. Hierro, J. M. Arizmendi, G. Montoya, A. Prado, and A. Muga. 2003. Activation mechanism of the nuclear chaperone nucleoplasmin: role of the core domain. J. Mol. Biol. 334:585-593. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources