

Eukaryotic translation initiation factor 3 (eIF3) and eIF2 can promote mRNA binding to 40S subunits independently of eIF4G in yeast
- PMID: 16449648
- PMCID: PMC1367198
- DOI: 10.1128/MCB.26.4.1355-1372.2006
Eukaryotic translation initiation factor 3 (eIF3) and eIF2 can promote mRNA binding to 40S subunits independently of eIF4G in yeast
Abstract
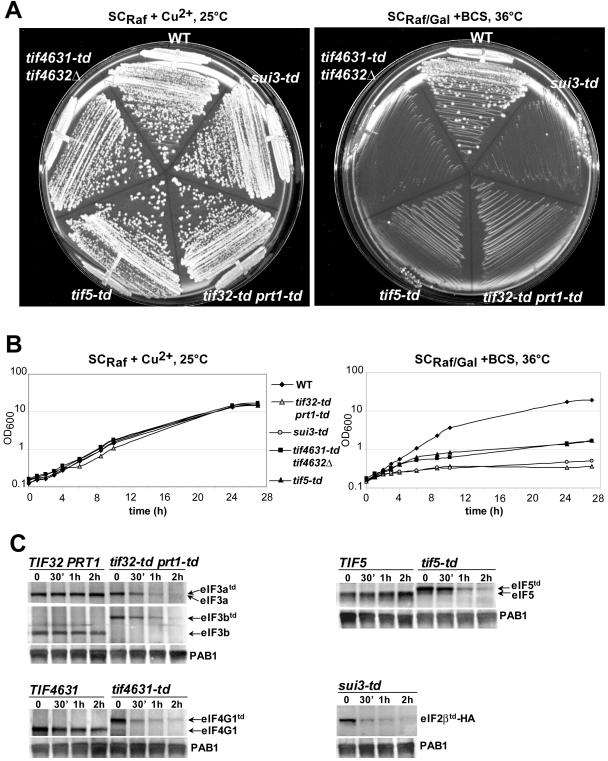
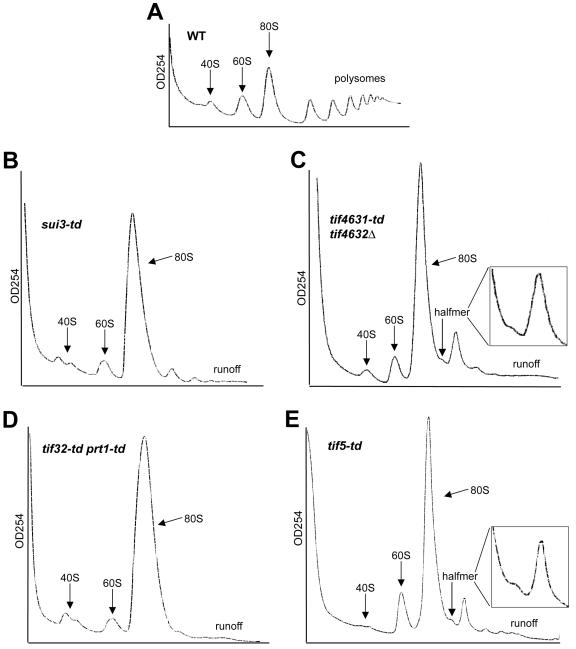
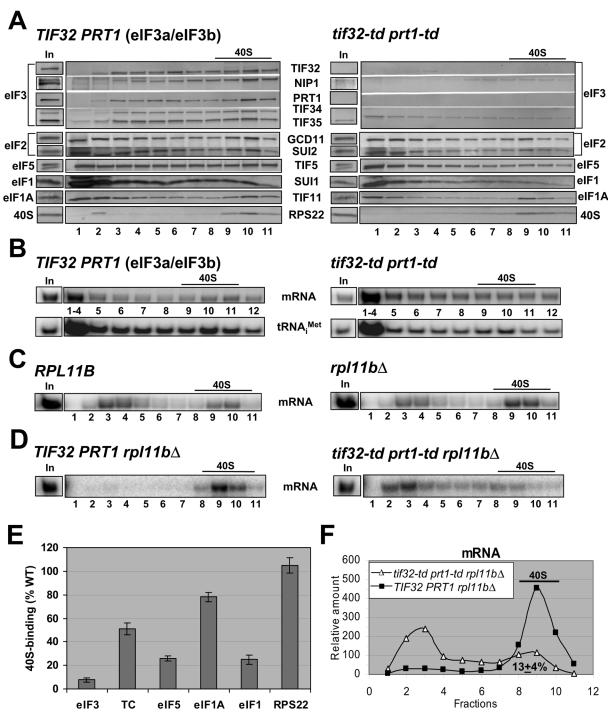
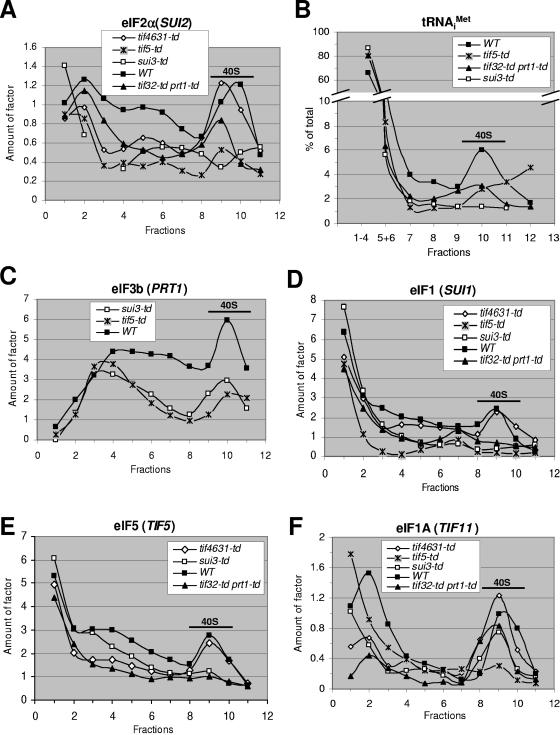
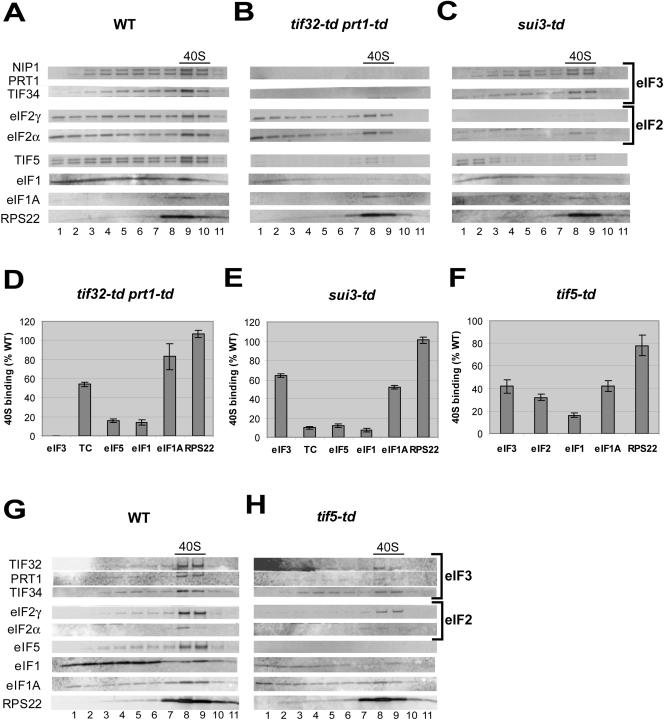
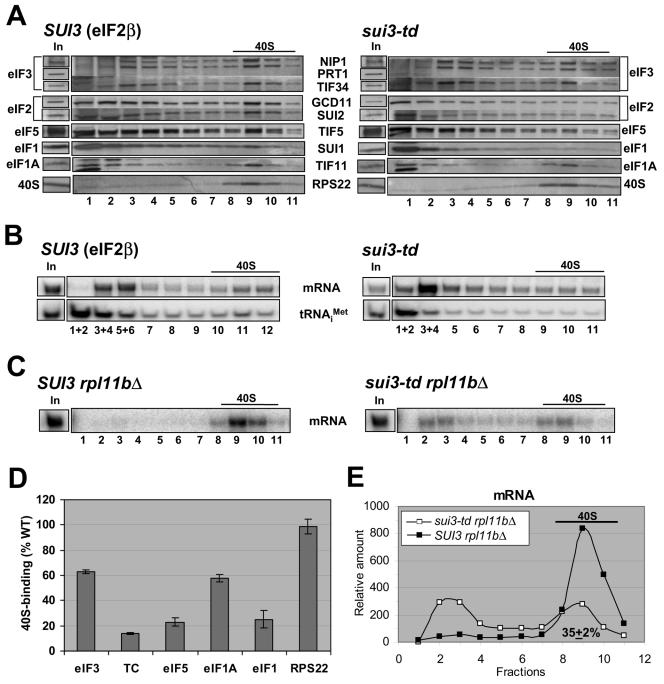
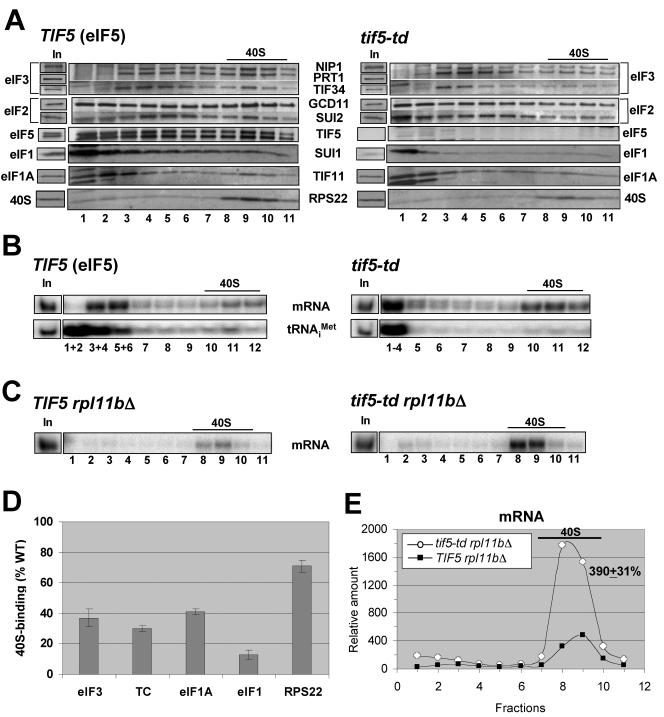
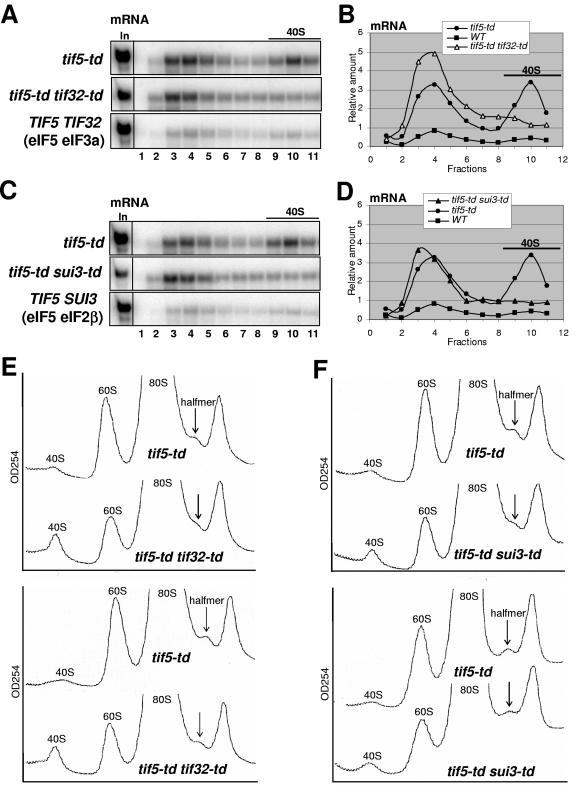
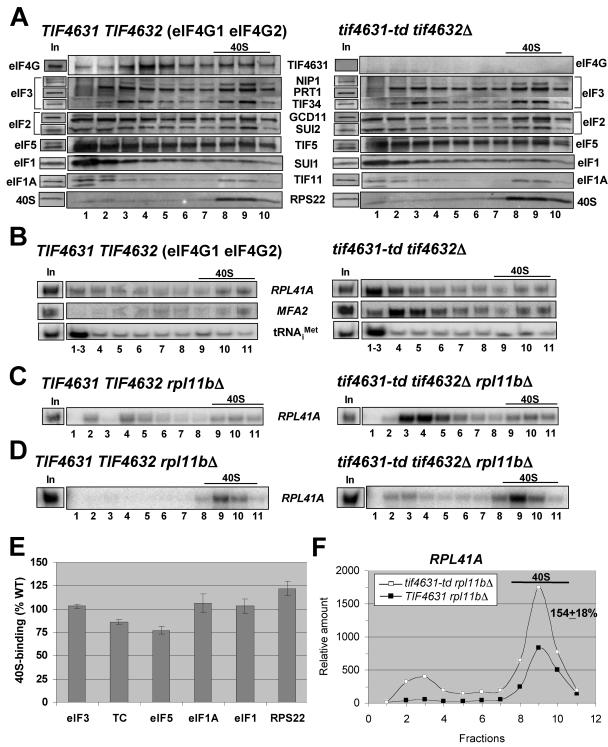
Recruitment of the eukaryotic translation initiation factor 2 (eIF2)-GTP-Met-tRNAiMet ternary complex to the 40S ribosome is stimulated by multiple initiation factors in vitro, including eIF3, eIF1, eIF5, and eIF1A. Recruitment of mRNA is thought to require the functions of eIF4F and eIF3, with the latter serving as an adaptor between the ribosome and the 4G subunit of eIF4F. To define the factor requirements for these reactions in vivo, we examined the effects of depleting eIF2, eIF3, eIF5, or eIF4G in Saccharomyces cerevisiae cells on binding of the ternary complex, other initiation factors, and RPL41A mRNA to native 43S and 48S preinitiation complexes. Depleting eIF2, eIF3, or eIF5 reduced 40S binding of all constituents of the multifactor complex (MFC), comprised of these three factors and eIF1, supporting a mechanism of coupled 40S binding by MFC components. 40S-bound mRNA strongly accumulated in eIF5-depleted cells, even though MFC binding to 40S subunits was reduced by eIF5 depletion. Hence, stimulation of the GTPase activity of the ternary complex, a prerequisite for 60S subunit joining in vitro, is likely the rate-limiting function of eIF5 in vivo. Depleting eIF2 or eIF3 impaired mRNA binding to free 40S subunits, but depleting eIF4G led unexpectedly to accumulation of mRNA on 40S subunits. Thus, it appears that eIF3 and eIF2 are more critically required than eIF4G for stable binding of at least some mRNAs to native preinitiation complexes and that eIF4G has a rate-limiting function at a step downstream of 48S complex assembly in vivo.
Figures
References
-
- Asano, K., T. G. Kinzy, W. C. Merrick, and J. W. B. Hershey. 1997. Conservation and diversity of eukaryotic translation initiation factor eIF3. J. Biol. Chem. 272:1101-1109. - PubMed
-
- Asano, K., L. Phan, L. Valasek, L. W. Schoenfeld, A. Shalev, J. Clayton, K. Nielsen, T. F. Donahue, and A. G. Hinnebusch. 2001. A multifactor complex of eIF1, eIF2, eIF3, eIF5, and tRNA(i)Met promotes initiation complex assembly and couples GTP hydrolysis to AUG recognition. Cold Spring Harbor Symp. Quant. Biol. 66:403-415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous