

Efficient and specific targeting of Polycomb group proteins requires cooperative interaction between Grainyhead and Pleiohomeotic
- PMID: 16449654
- PMCID: PMC1367177
- DOI: 10.1128/MCB.26.4.1434-1444.2006
Efficient and specific targeting of Polycomb group proteins requires cooperative interaction between Grainyhead and Pleiohomeotic
Abstract
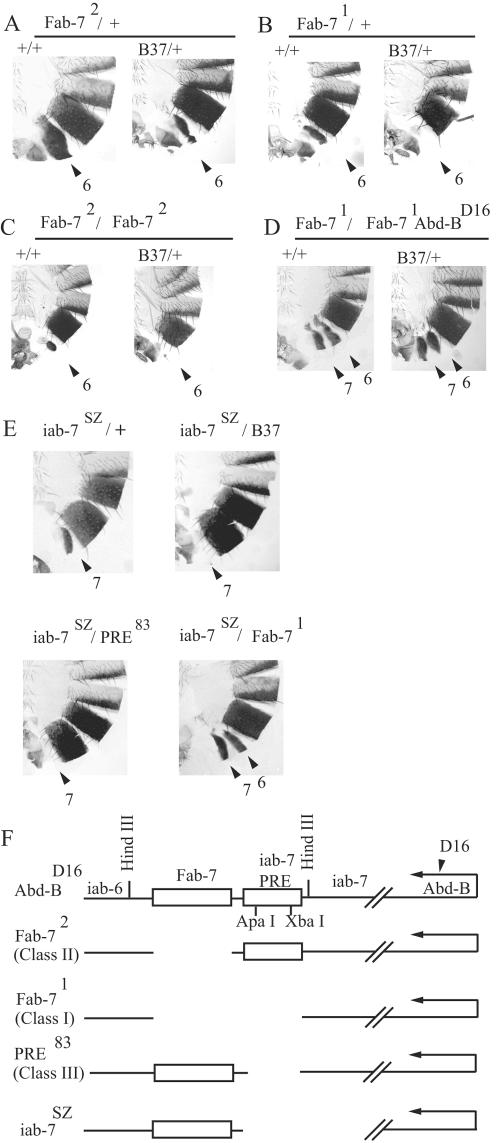
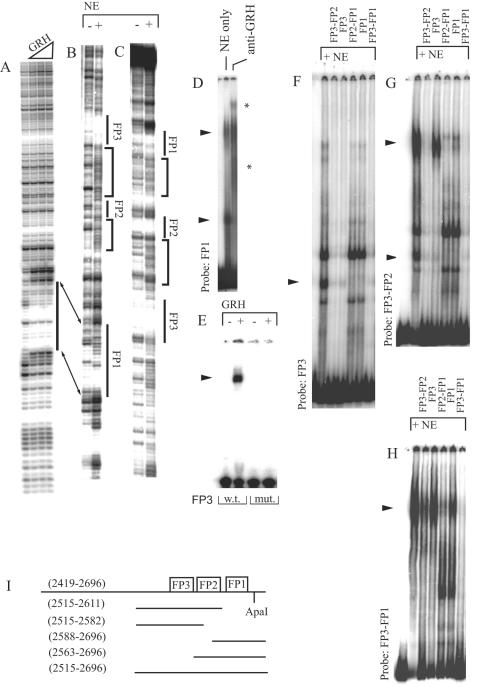
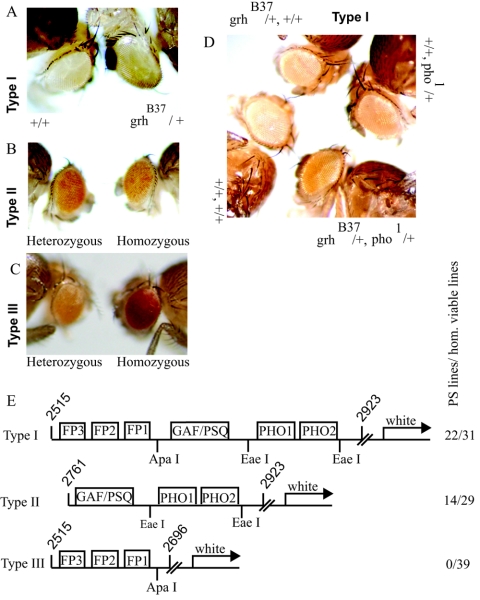
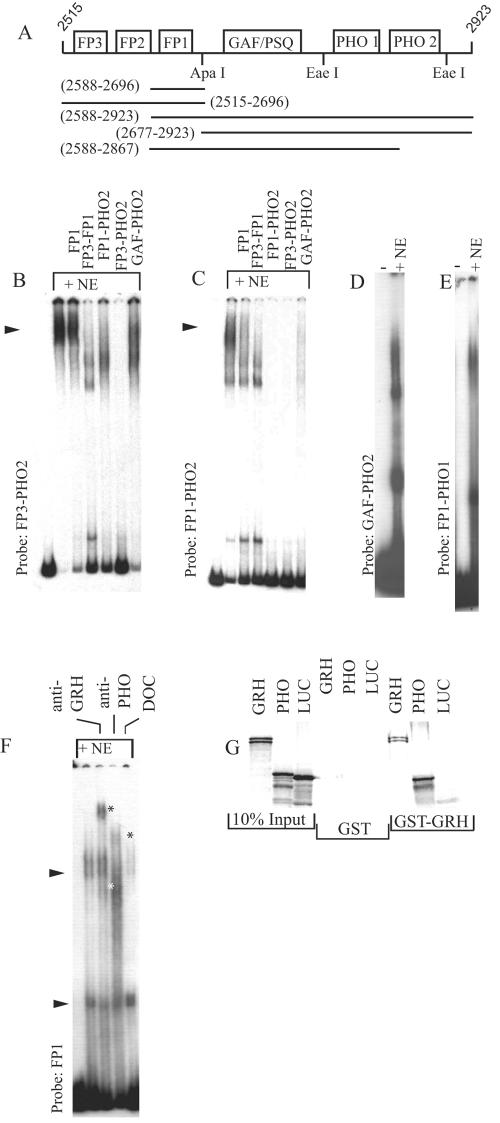
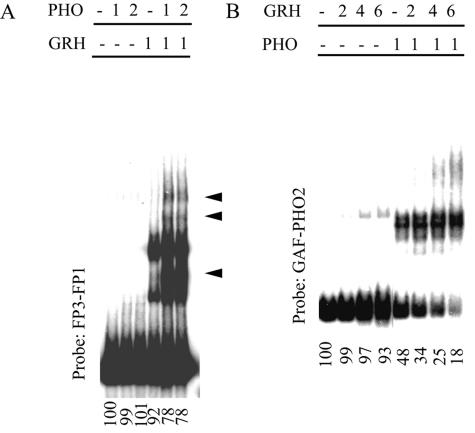
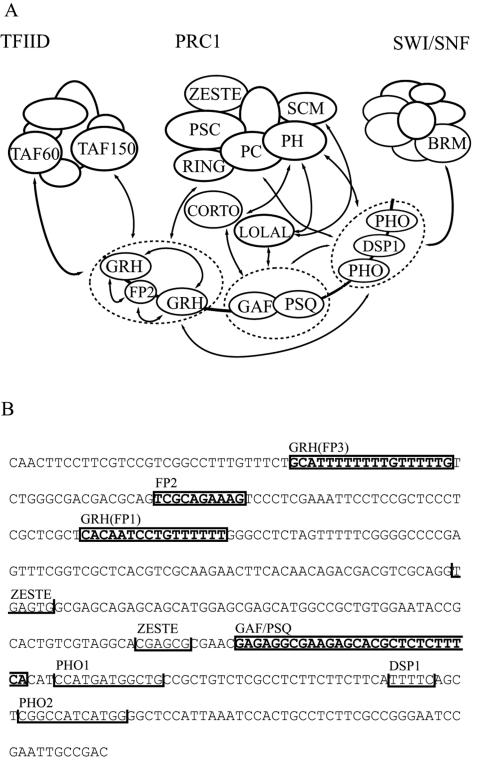
Specific targeting of the protein complexes formed by the Polycomb group of proteins is critically required to maintain the inactive state of a group of developmentally regulated genes. Although the role of DNA binding proteins in this process has been well established, it is still not understood how these proteins target the Polycomb complexes specifically to their response elements. Here we show that the grainyhead gene, which encodes a DNA binding protein, interacts with one such Polycomb response element of the bithorax complex. Grainyhead binds to this element in vitro. Moreover, grainyhead interacts genetically with pleiohomeotic in a transgene-based, pairing-dependent silencing assay. Grainyhead also interacts with Pleiohomeotic in vitro, which facilitates the binding of both proteins to their respective target DNAs. Such interactions between two DNA binding proteins could provide the basis for the cooperative assembly of a nucleoprotein complex formed in vitro. Based on these results and the available data, we propose that the role of DNA binding proteins in Polycomb group-dependent silencing could be described by a model very similar to that of an enhanceosome, wherein the unique arrangement of protein-protein interaction modules exposed by the cooperatively interacting DNA binding proteins provides targeting specificity.
Figures

References
-
- Attardi, L. D., and R. Tjian. 1993. Drosophila tissue-specific transcription factor NTF-1 contains a novel isoleucine-rich activation motif. Genes Dev. 7:1341-1353. - PubMed
-
- Bray, S. J., B. Burke, N. H. Brown, and J. Hirsh. 1989. Embryonic expression pattern of a family of Drosophila proteins that interact with a central nervous system regulatory element. Genes Dev. 3:1130-1145. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases